

Using RDNA sequences to define dinoflagellate species
- PMID: 35213572
- PMCID: PMC8880924
- DOI: 10.1371/journal.pone.0264143
Using RDNA sequences to define dinoflagellate species
Abstract
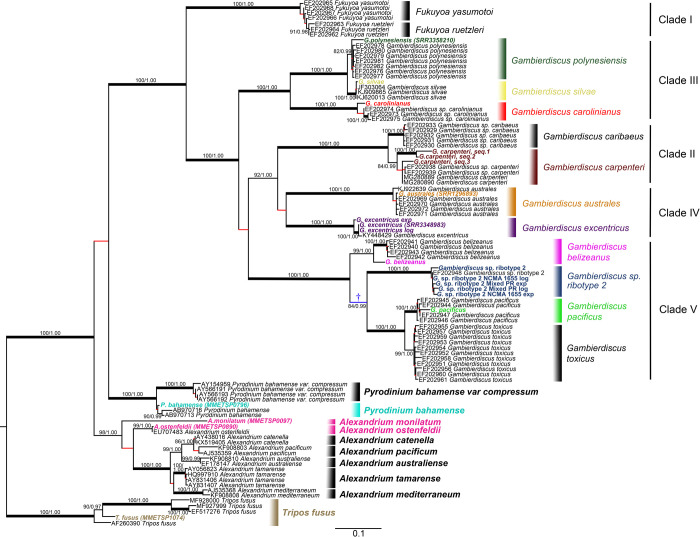
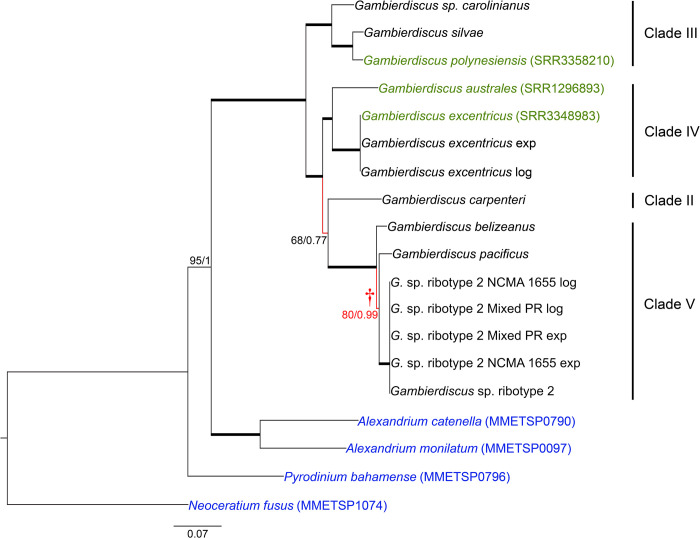
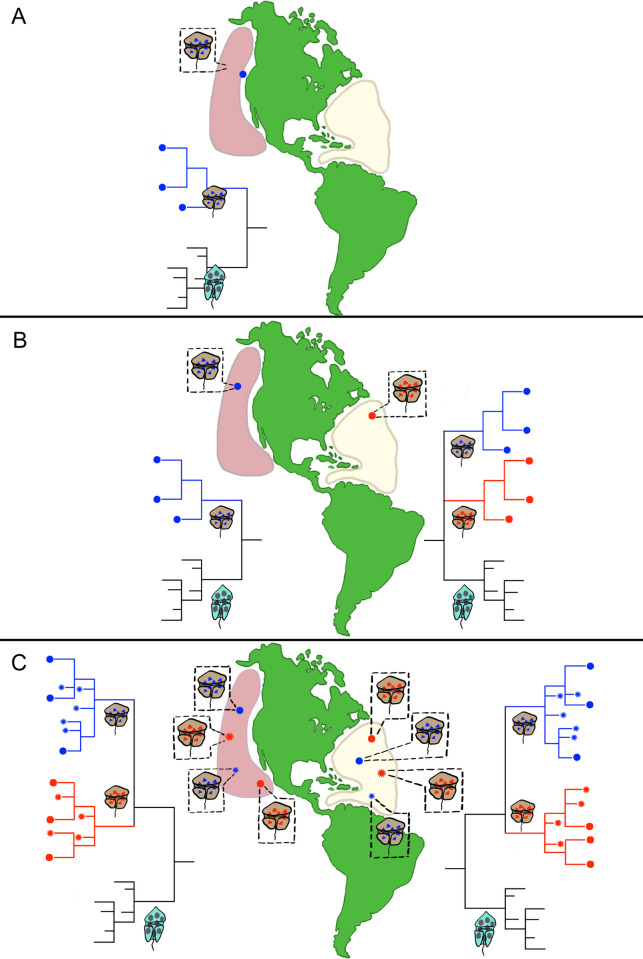
Dinoflagellate species are traditionally defined using morphological characters, but molecular evidence accumulated over the past several decades indicates many morphologically-based descriptions are inaccurate. This recognition led to an increasing reliance on DNA sequence data, particularly rDNA gene segments, in defining species. The validity of this approach assumes the divergence in rDNA or other selected genes parallels speciation events. Another concern is whether single gene rDNA phylogenies by themselves are adequate for delineating species or if multigene phylogenies are required instead. Currently, few studies have directly assessed the relative utility of multigene versus rDNA-based phylogenies for distinguishing species. To address this, the current study examined D1-D3 and ITS/5.8S rDNA gene regions, a multi-gene phylogeny, and morphological characters in Gambierdiscus and other related dinoflagellate genera to determine if they produce congruent phylogenies and identify the same species. Data for the analyses were obtained from previous sequencing efforts and publicly available dinoflagellate transcriptomic libraries as well from the additional nine well-characterized Gambierdiscus species transcriptomic libraries generated in this study. The D1-D3 and ITS/5.8S phylogenies successfully identified the described Gambierdiscus and Alexandrium species. Additionally, the data showed that the D1-D3 and multigene phylogenies were equally capable of identifying the same species. The multigene phylogenies, however, showed different relationships among species and are likely to prove more accurate at determining phylogenetic relationships above the species level. These data indicated that D1-D3 and ITS/5.8S rDNA region phylogenies are generally successful for identifying species of Gambierdiscus, and likely those of other dinoflagellates. To assess how broadly general this finding is likely to be, rDNA molecular phylogenies from over 473 manuscripts representing 232 genera and 863 described species of dinoflagellates were reviewed. Results showed the D1-D3 rDNA and ITS phylogenies in combination are capable of identifying 97% of dinoflagellate species including all the species belonging to the genera Alexandrium, Ostreopsis and Gambierdiscus, although it should be noted that multi-gene phylogenies are preferred for inferring relationships among these species. A protocol is presented for determining when D1-D3, confirmed by ITS/5.8S rDNA sequence data, would take precedence over morphological features when describing new dinoflagellate species. This protocol addresses situations such as: a) when a new species is both morphologically and molecularly distinct from other known species; b) when a new species and closely related species are morphologically indistinguishable, but genetically distinct; and c) how to handle potentially cryptic species and cases where morphotypes are clearly distinct but have the same rDNA sequence. The protocol also addresses other molecular, morphological, and genetic approaches required to resolve species boundaries in the small minority of species where the D1-D3/ITS region phylogenies fail.
Conflict of interest statement
The authors have read the journal’s policy and have the following competing interests: R. Wayne Litaker is a paid contractor for the National Oceanic and Atmospheric Administration (NOAA).
Figures



References
-
- Stoecker DK. Mixotrophy among Dinoflagellates. J Eukaryot Microbiol. 1999;46: 397–401. doi: 10.1111/j.1550-7408.1999.tb04619.x - DOI
-
- Taylor FJR, Hoppenrath M, Saldarriaga JF. Dinoflagellate diversity and distribution. Biodivers Conserv. 2008;17: 407–418. doi: 10.1007/s10531-007-9258-3 - DOI
-
- Jeong HJ, Yoo YD, Kim JS, Seong KA, Kang NS, Kim TH. Growth, feeding and ecological roles of the mixotrophic and heterotrophic dinoflagellates in marine planktonic food webs. Ocean Sci J. 2010;45: 65–91. doi: 10.1007/s12601-010-0007-2 - DOI
-
- Gómez F. A quantitative review of the lifestyle, habitat and trophic diversity of dinoflagellates (Dinoflagellata, Alveolata). Syst Biodivers. 2012;10: 267–275. doi: 10.1080/14772000.2012.721021 - DOI
-
- Litaker RW, Vandersea MW, Kibler SR, Madden VJ, Noga EJ, Tester PA. Life Cycle of the Heterotrophic Dinoflagellate Pfiesteria piscicida (Dinophyceae). J Phycol. 2002;38: 442–463. doi: 10.1046/j.1529-8817.2002.01242.x - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
