

Immunization with GP1 but Not Core-like Particles Displaying Isolated Receptor-Binding Epitopes Elicits Virus-Neutralizing Antibodies against Junín Virus
- PMID: 35214632
- PMCID: PMC8874384
- DOI: 10.3390/vaccines10020173
Immunization with GP1 but Not Core-like Particles Displaying Isolated Receptor-Binding Epitopes Elicits Virus-Neutralizing Antibodies against Junín Virus
Abstract
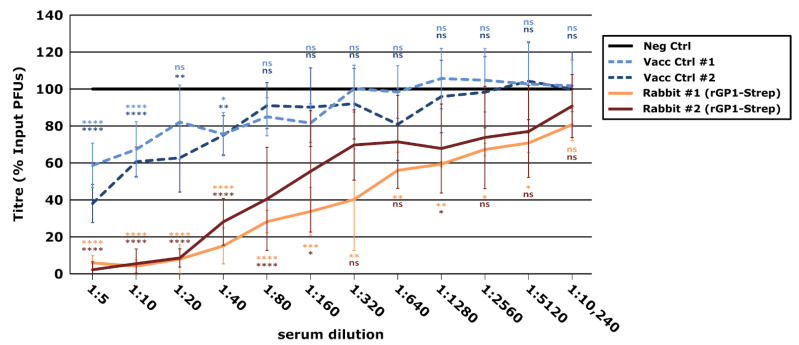
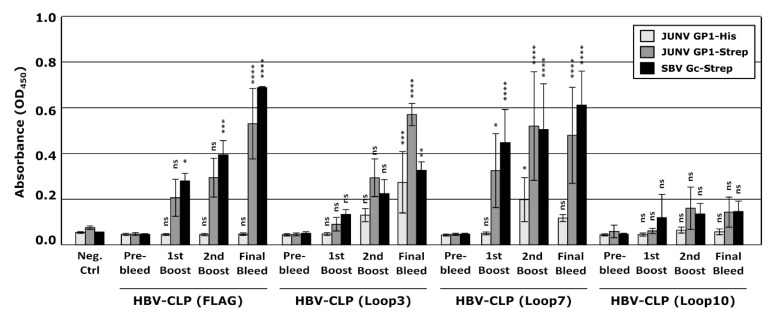
New World arenaviruses are rodent-transmitted viruses and include a number of pathogens that are responsible for causing severe human disease. This includes Junín virus (JUNV), which is the causative agent of Argentine hemorrhagic fever. The wild nature and mobility of the rodent reservoir host makes it difficult to control the disease, and currently passive immunization with high-titer neutralizing antibody-containing plasma from convalescent patients is the only specific therapy. However, dwindling supplies of naturally available convalescent plasma, and challenges in developing similar resources for other closely related viruses, have made the development of alternative antibody-based therapeutic approaches of critical importance. In this study, we sought to induce a neutralizing antibody response in rabbits against the receptor-binding subunit of the viral glycoprotein, GP1, and the specific peptide sequences in GP1 involved in cellular receptor contacts. While these specific receptor-interacting peptides did not efficiently induce the production of neutralizing antibodies when delivered as a particulate antigen (as part of hepatitis B virus core-like particles), we showed that recombinant JUNV GP1 purified from transfected mammalian cells induced virus-neutralizing antibodies at high titers in rabbits. Further, neutralization was observed across a range of unrelated JUNV strains, a feature that is critical for effectiveness in the field. These results underscore the potential of GP1 alone to induce a potent neutralizing antibody response and highlight the importance of epitope presentation. In addition, effective virus neutralization by rabbit antibodies supports the potential applicability of this species for the future development of immunotherapeutics (e.g., based on humanized monoclonal antibodies). Such information can be applied in the design of vaccines and immunogens for both prevention and specific therapies against this and likely also other closely related pathogenic New World arenaviruses.
Keywords: GP1; Junín virus; arenavirus; glycoprotein; immune response; neutralizing antibodies.
Conflict of interest statement
The authors declare no conflict of interest. The sponsors had no role in the design, execution, interpretation, or writing of the study.
Figures
References
-
- Mills J.N., Ellis B.A., Childs J.E., McKee K.T., Maiztegui J.I., Peters C.J., Ksiazek T.G., Jahrling P.B. Prevalence of infection with Junin virus in rodent populations in the epidemic area of Argentine hemorrhagic fever. Am. J. Trop. Med. Hyg. 1994;51:554–562. doi: 10.4269/ajtmh.1994.51.554. - DOI - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
