

What Is the Evidence That Dietary Macronutrient Composition Influences Exercise Performance? A Narrative Review
- PMID: 35215511
- PMCID: PMC8875928
- DOI: 10.3390/nu14040862
What Is the Evidence That Dietary Macronutrient Composition Influences Exercise Performance? A Narrative Review
Abstract
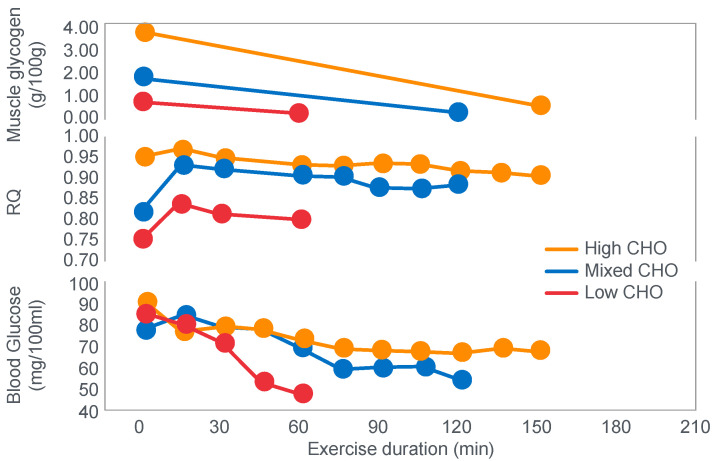
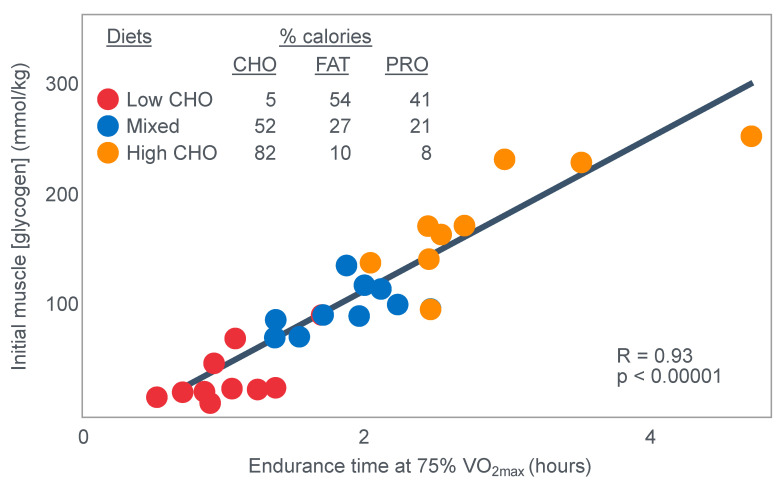
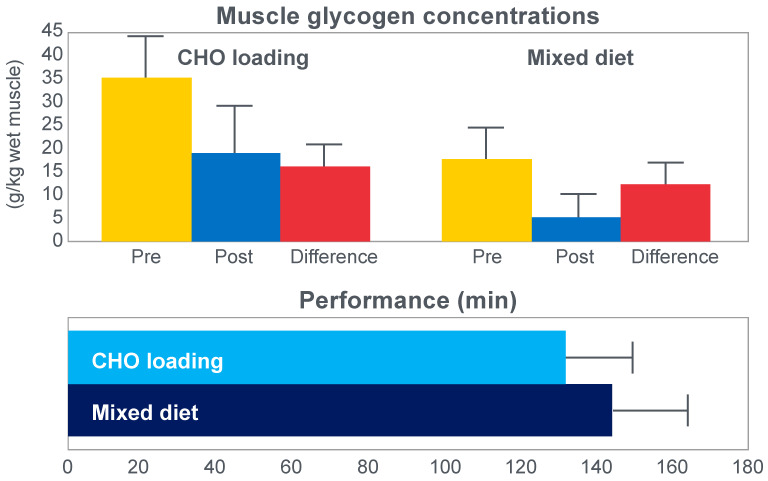
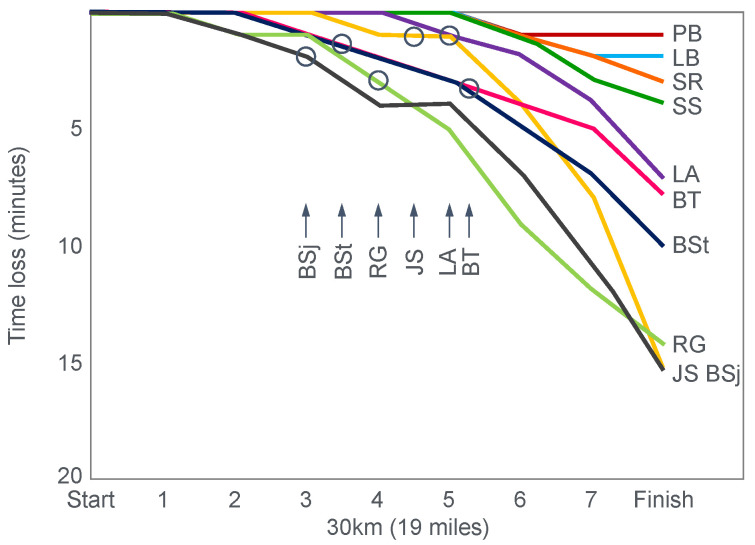
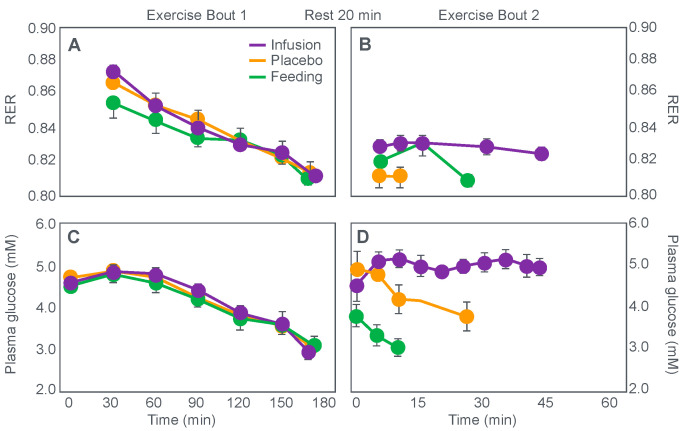
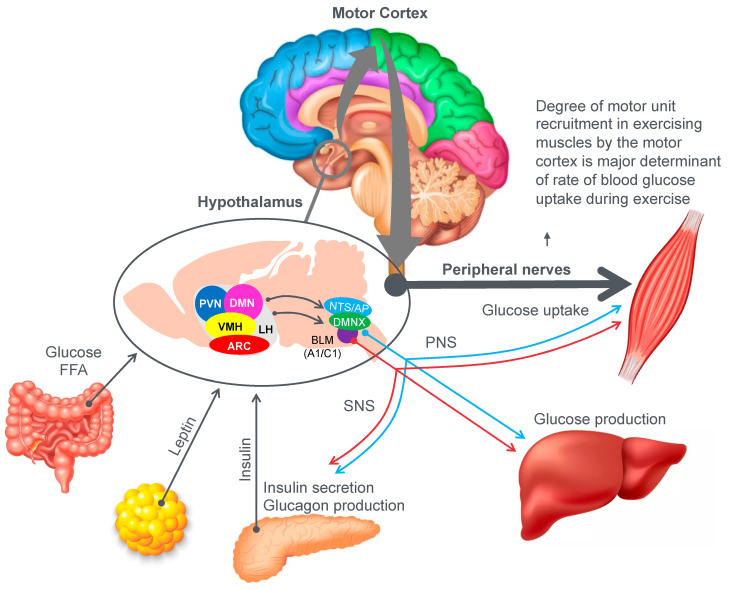
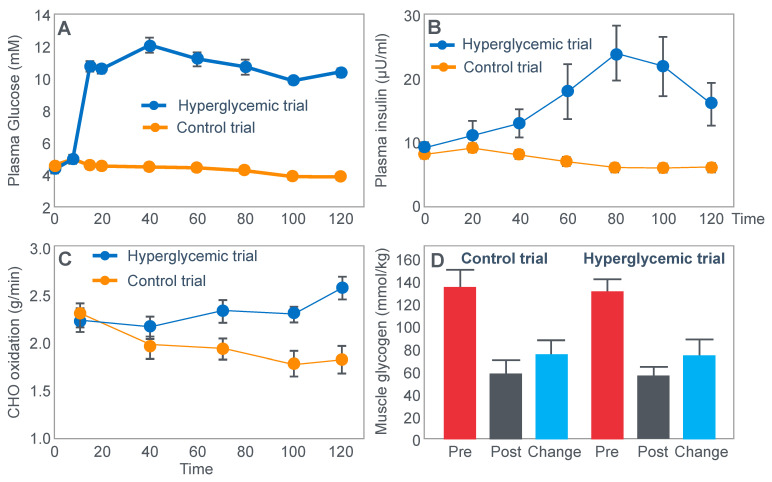
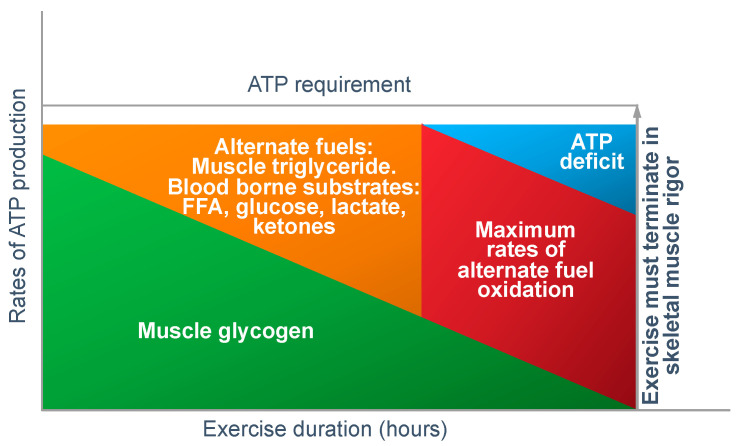
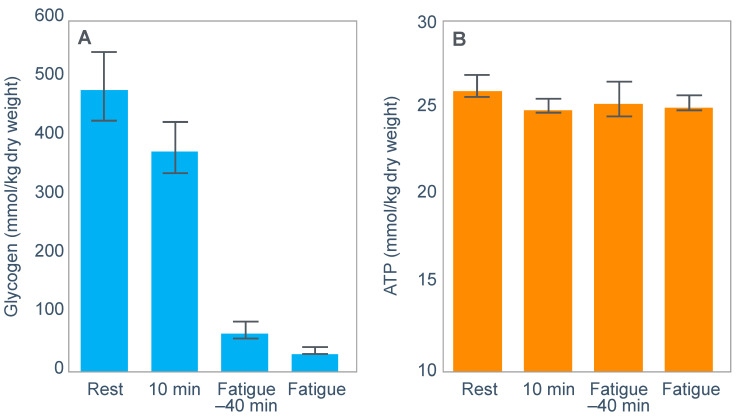
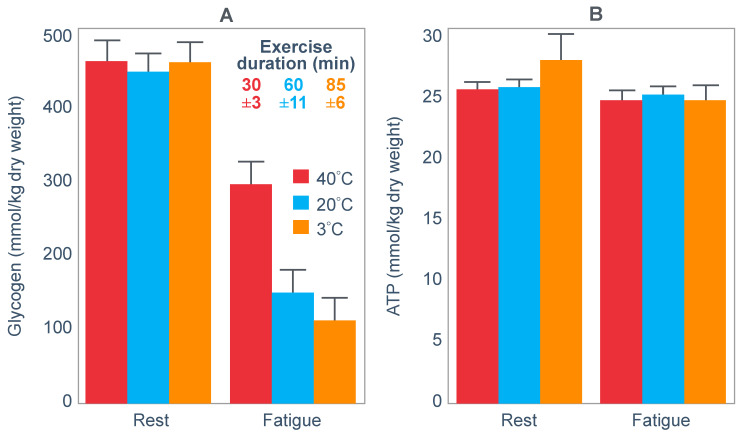
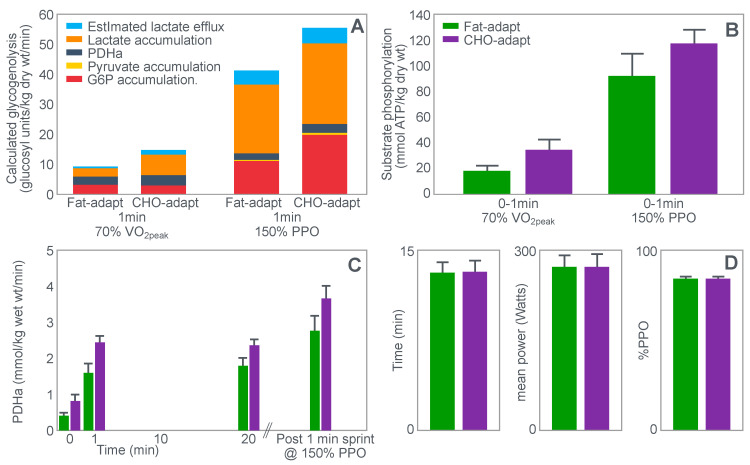
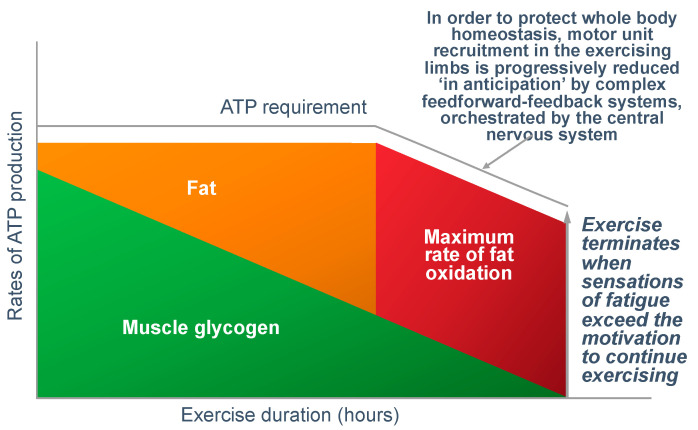
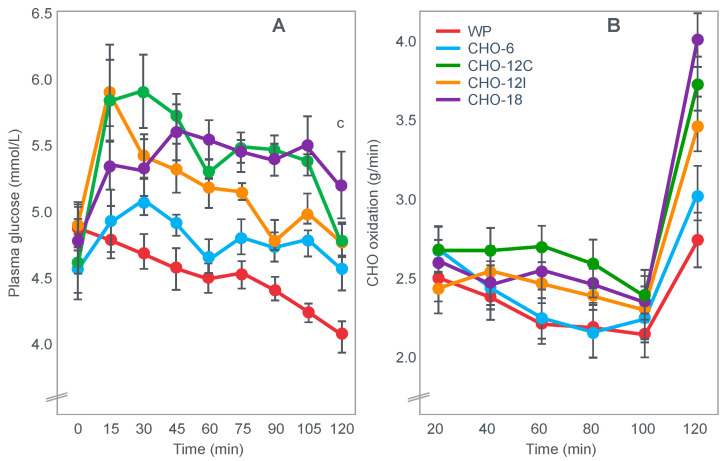
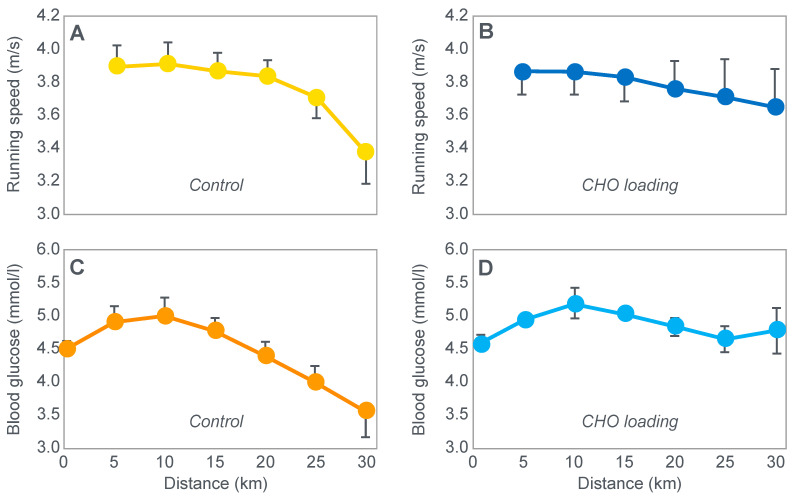
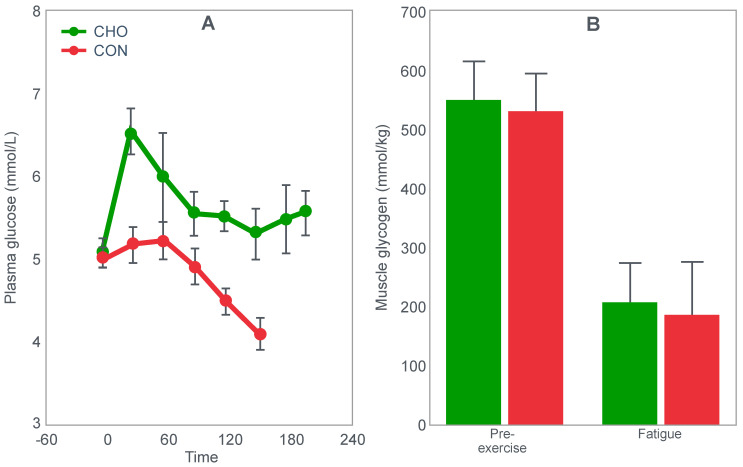
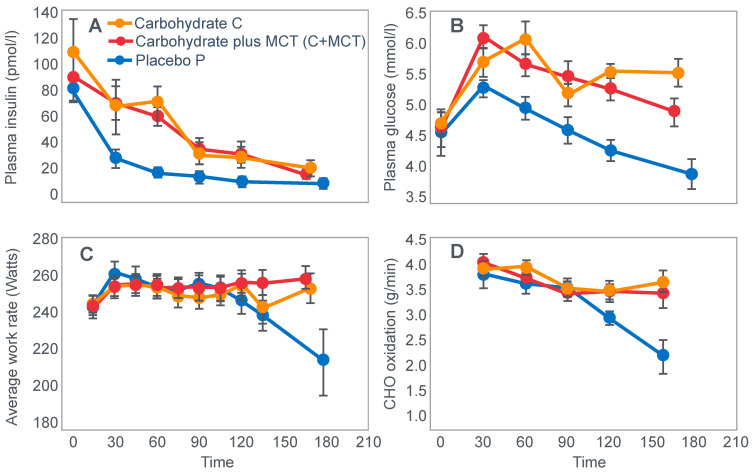
The introduction of the needle muscle biopsy technique in the 1960s allowed muscle tissue to be sampled from exercising humans for the first time. The finding that muscle glycogen content reached low levels at exhaustion suggested that the metabolic cause of fatigue during prolonged exercise had been discovered. A special pre-exercise diet that maximized pre-exercise muscle glycogen storage also increased time to fatigue during prolonged exercise. The logical conclusion was that the athlete's pre-exercise muscle glycogen content is the single most important acutely modifiable determinant of endurance capacity. Muscle biochemists proposed that skeletal muscle has an obligatory dependence on high rates of muscle glycogen/carbohydrate oxidation, especially during high intensity or prolonged exercise. Without this obligatory carbohydrate oxidation from muscle glycogen, optimum muscle metabolism cannot be sustained; fatigue develops and exercise performance is impaired. As plausible as this explanation may appear, it has never been proven. Here, I propose an alternate explanation. All the original studies overlooked one crucial finding, specifically that not only were muscle glycogen concentrations low at exhaustion in all trials, but hypoglycemia was also always present. Here, I provide the historical and modern evidence showing that the blood glucose concentration-reflecting the liver glycogen rather than the muscle glycogen content-is the homeostatically-regulated (protected) variable that drives the metabolic response to prolonged exercise. If this is so, nutritional interventions that enhance exercise performance, especially during prolonged exercise, will be those that assist the body in its efforts to maintain the blood glucose concentration within the normal range.
Keywords: carbohydrates; diet; endurance; fatigue; fats; hypoglycaemia; liver glycogen; muscle glycogen.
Conflict of interest statement
The author declares that he has no conflict of interest. Although he writes and has written books promoting the low-carbohydrate diet, all proceeds from those publications are donated to his Non-Profit Organization, The Noakes Foundation. In the past, he received funding from commercial companies to research the effects of carbohydrate ingestion and high-carbohydrate diets on athletic performance, which ceased more than 15 years ago. He did not personally benefit financially from such funding. Author royalties from his other books, Lore of Running (2002) and Waterlogged: The Serious Problem of Overhydration in Endurance Sports (2011), are donated to the Tim and Marilyn Sports Science Research Trust that funds a post-doctoral scholarship for academic work in sport and health promotion.
Figures
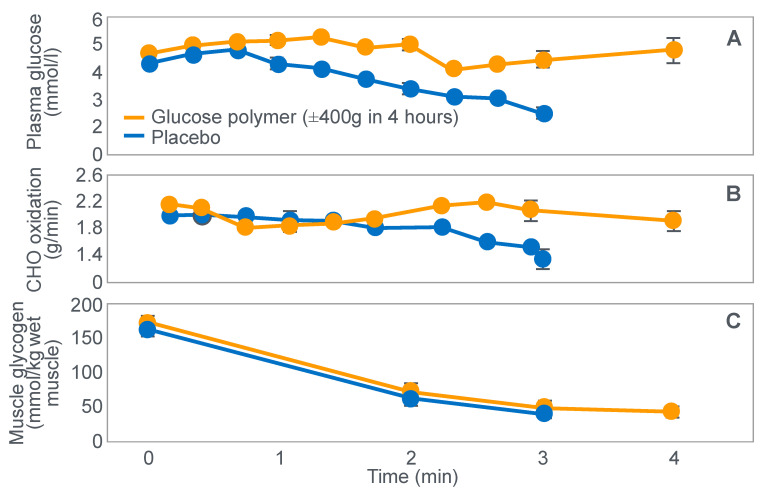
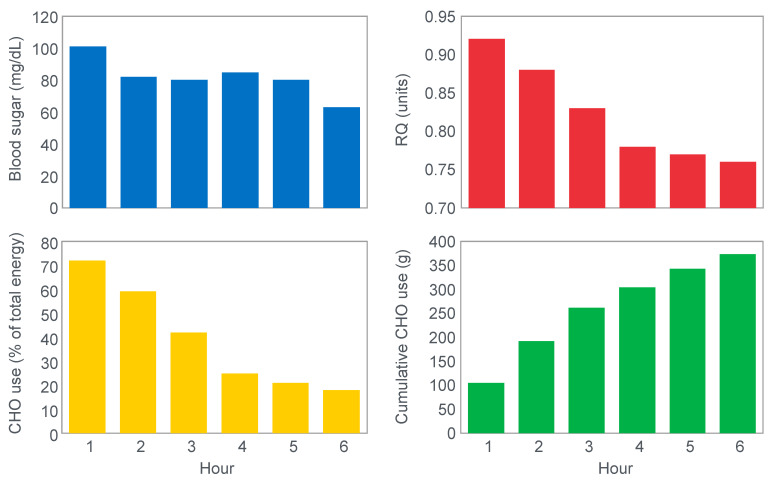
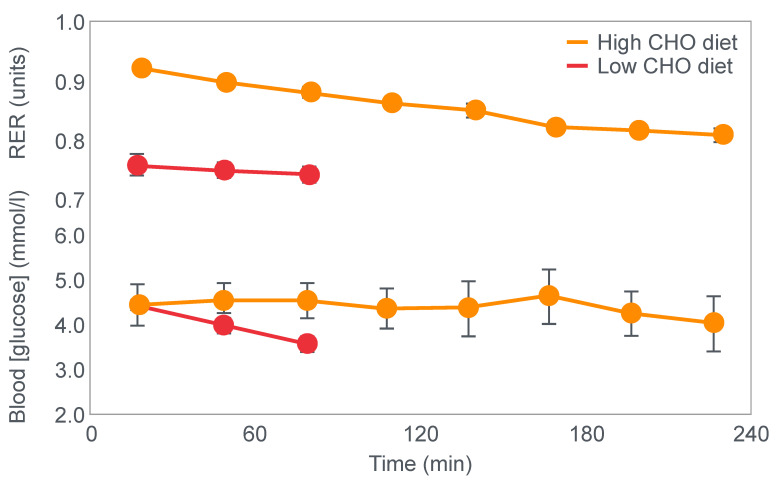
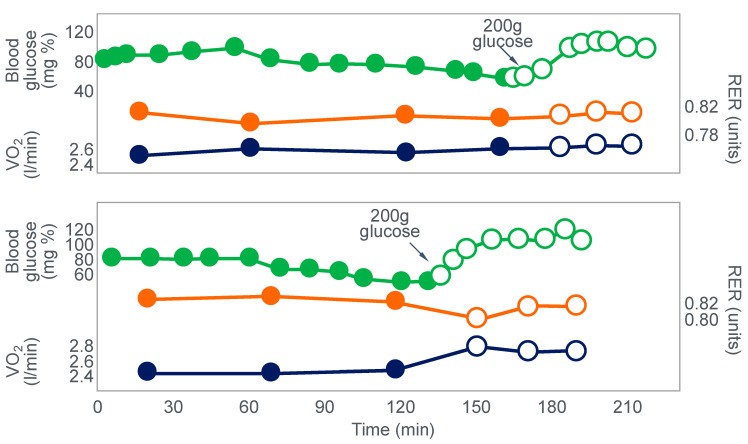
References
-
- Cathcart E.P. The influence of muscle work on protein metabolism. Physiol. Rev. 1925;5:225–243. doi: 10.1152/physrev.1925.5.2.225. - DOI
-
- Hill A.V., Long C.H.N., Lupton H. Muscular exercise, lactic acid and the supply and utilisation of oxygen: Parts VII–VIII. Proc. R. Soc. Lond. B Biol. Sci. 1924;97:155–176.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
