

A Novel Motif in the 3'-UTR of PRRSV-2 Is Critical for Viral Multiplication and Contributes to Enhanced Replication Ability of Highly Pathogenic or L1 PRRSV
- PMID: 35215760
- PMCID: PMC8875199
- DOI: 10.3390/v14020166
A Novel Motif in the 3'-UTR of PRRSV-2 Is Critical for Viral Multiplication and Contributes to Enhanced Replication Ability of Highly Pathogenic or L1 PRRSV
Abstract
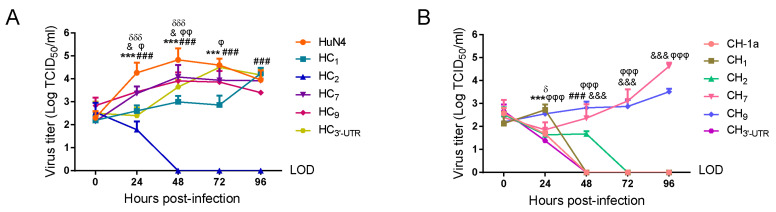
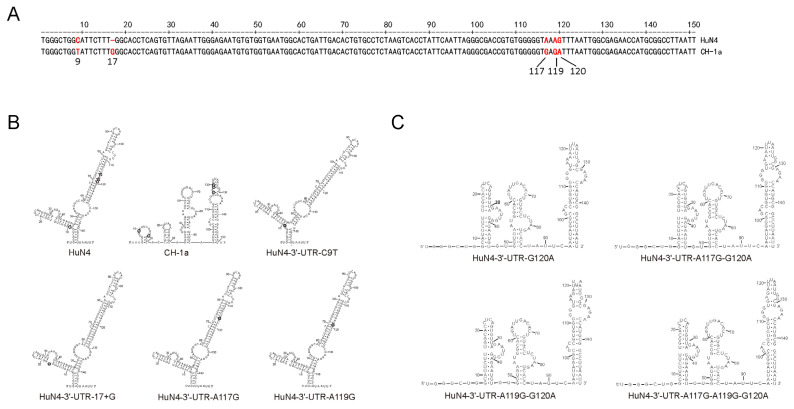
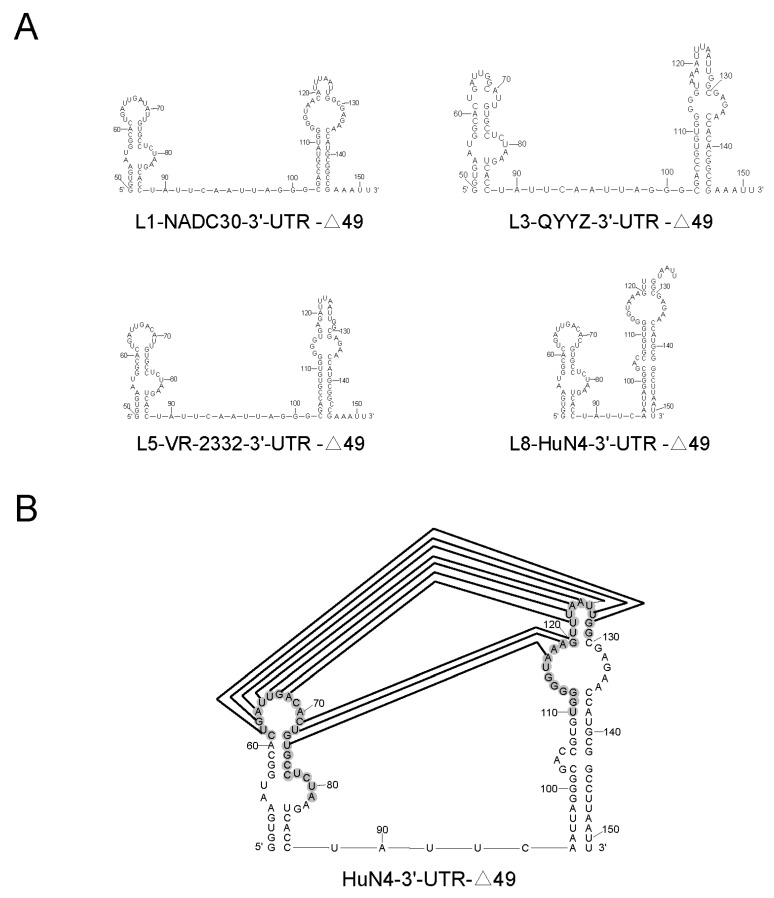
Highly pathogenic porcine reproductive and respiratory syndrome virus (HP-PRRSV) with enhanced replication capability emerged in China and has become dominant epidemic strain since 2006. Up to now, the replication-regulated genes of PRRSV have not been fully clarified. Here, by swapping the genes or elements between HP-PRRSV and classical PRRSV based on infectious clones, NSP1, NSP2, NSP7, NSP9 and 3'-UTR are found to contribute to the high replication efficiency of HP-PRRSV. Further study revealed that mutations at positions 117th or 119th in the 3'-UTR are significantly related to replication efficiency, and the nucleotide at position 120th is critical for viral rescue. The motif composed by 117-120th nucleotides was quite conservative within each lineage of PRRSV; mutations in the motif of HP-PRRSV and currently epidemic lineage 1 (L1) PRRSV showed higher synthesis ability of viral negative genomic RNA, suggesting that those mutations were beneficial for viral replication. RNA structure analysis revealed that this motif maybe involved into a pseudoknot in the 3'-UTR. The results discovered a novel motif, 117-120th nucleotide in the 3'-UTR, that is critical for replication of PRRSV-2, and mutations in the motif contribute to the enhanced replicative ability of HP-PRRSV or L1 PRRSV. Our findings will help to understand the molecular basis of PRRSV replication and find the potential factors resulting in an epidemic strain of PRRSV.
Keywords: 3′-UTR; HP-PRRSV; epidemic; motif; replication; secondary structure.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures

Similar articles
-
Two Residues in NSP9 Contribute to the Enhanced Replication and Pathogenicity of Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus.J Virol. 2018 Mar 14;92(7):e02209-17. doi: 10.1128/JVI.02209-17. Print 2018 Apr 1. J Virol. 2018. PMID: 29321316 Free PMC article.
-
Genomic sequencing reveals mutations potentially related to the overattenuation of a highly pathogenic porcine reproductive and respiratory syndrome virus.Clin Vaccine Immunol. 2013 Apr;20(4):613-9. doi: 10.1128/CVI.00672-12. Epub 2013 Feb 13. Clin Vaccine Immunol. 2013. PMID: 23408525 Free PMC article.
-
Nonstructural protein 9 residues 586 and 592 are critical sites in determining the replication efficiency and fatal virulence of the Chinese highly pathogenic porcine reproductive and respiratory syndrome virus.Virology. 2018 Apr;517:135-147. doi: 10.1016/j.virol.2018.01.018. Epub 2018 Feb 12. Virology. 2018. PMID: 29397202 Free PMC article.
-
Identification of dispensable nucleotide sequence in 3' untranslated region of porcine reproductive and respiratory syndrome virus.Virus Res. 2010 Dec;154(1-2):38-47. doi: 10.1016/j.virusres.2010.08.027. Epub 2010 Sep 15. Virus Res. 2010. PMID: 20833212 Free PMC article. Review.
-
Non-structural protein 2 of the porcine reproductive and respiratory syndrome (PRRS) virus: a crucial protein in viral pathogenesis, immunity and diagnosis.Res Vet Sci. 2013 Aug;95(1):1-7. doi: 10.1016/j.rvsc.2013.03.015. Epub 2013 Apr 13. Res Vet Sci. 2013. PMID: 23591056 Review.
Cited by
-
Case report: Clinical and virological characteristics of aseptic meningitis caused by a recombinant echovirus 18 in an immunocompetent adult.Front Med (Lausanne). 2023 Jan 12;9:1094347. doi: 10.3389/fmed.2022.1094347. eCollection 2022. Front Med (Lausanne). 2023. PMID: 36714132 Free PMC article.
-
Early Highly Pathogenic Porcine Reproductive and Respiratory Syndrome Virus Infection Induces Necroptosis in Immune Cells of Peripheral Lymphoid Organs.Viruses. 2025 Feb 20;17(3):290. doi: 10.3390/v17030290. Viruses. 2025. PMID: 40143222 Free PMC article.
-
Reverse genetics construction and pathogenicity of a novel recombinant NADC30-like PRRSV isolated in China.Front Vet Sci. 2024 Jun 26;11:1434539. doi: 10.3389/fvets.2024.1434539. eCollection 2024. Front Vet Sci. 2024. PMID: 38993278 Free PMC article.
-
Genomic characteristics of a novel emerging PRRSV branch in sublineage 8.7 in China.Front Microbiol. 2023 May 31;14:1186322. doi: 10.3389/fmicb.2023.1186322. eCollection 2023. Front Microbiol. 2023. PMID: 37323894 Free PMC article.
-
Research Progress on Porcine Reproductive and Respiratory Syndrome Virus NSP7 Protein.Animals (Basel). 2023 Jul 11;13(14):2269. doi: 10.3390/ani13142269. Animals (Basel). 2023. PMID: 37508047 Free PMC article. Review.
References
-
- Neumann E.J., Kliebenstein J.B., Johnson C.D., Mabry J.W., Bush E.J., Seitzinger A.H., Green A.L., Zimmerman J.J. Assessment of the economic impact of porcine reproductive and respiratory syndrome on swine production in the United States. J. Am. Vet. Med. Assoc. 2005;227:385–392. doi: 10.2460/javma.2005.227.385. - DOI - PubMed
-
- Wensvoort G., de Kluyver E.P., Pol J.M., Wagenaar F., Moormann R.J., Hulst M.M., Bloemraad R., den Besten A., Zetstra T., Terpstra C. Lelystad virus, the cause of porcine epidemic abortion and respiratory syndrome: A review of mystery swine disease research at Lelystad. Vet. Microbiol. 1992;33:185–193. doi: 10.1016/0378-1135(92)90046-V. - DOI - PubMed
-
- Gao J.C., Xiong J.Y., Ye C., Chang X.B., Guo J.C., Jiang C.G., Zhang G.H., Tian Z.J., Cai X.H., Tong G.Z., et al. Genotypic and geographical distribution of porcine reproductive and respiratory syndrome viruses in mainland China in 1996–2016. Vet. Microbiol. 2017;208:164–172. doi: 10.1016/j.vetmic.2017.08.003. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
