

Dengue and Zika Virus Capsid Proteins Contain a Common PEX19-Binding Motif
- PMID: 35215846
- PMCID: PMC8874546
- DOI: 10.3390/v14020253
Dengue and Zika Virus Capsid Proteins Contain a Common PEX19-Binding Motif
Abstract
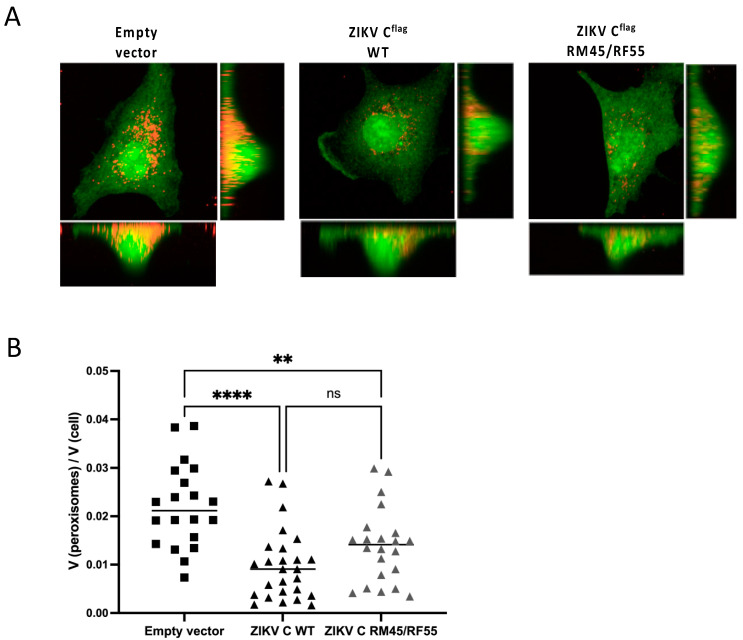
Flaviviruses such as dengue virus (DENV) and Zika virus (ZIKV) have evolved sophisticated mechanisms to suppress the host immune system. For instance, flavivirus infections were found to sabotage peroxisomes, organelles with an important role in innate immunity. The current model suggests that the capsid (C) proteins of DENV and ZIKV downregulate peroxisomes, ultimately resulting in reduced production of interferons by interacting with the host protein PEX19, a crucial chaperone in peroxisomal biogenesis. Here, we aimed to explore the importance of peroxisomes and the role of C interaction with PEX19 in the flavivirus life cycle. By infecting cells lacking peroxisomes we show that this organelle is required for optimal DENV replication. Moreover, we demonstrate that DENV and ZIKV C bind PEX19 through a conserved PEX19-binding motif, which is also commonly found in cellular peroxisomal membrane proteins (PMPs). However, in contrast to PMPs, this interaction does not result in the targeting of C to peroxisomes. Furthermore, we show that the presence of C results in peroxisome loss due to impaired peroxisomal biogenesis, which appears to occur by a PEX19-independent mechanism. Hence, these findings challenge the current model of how flavivirus C might downregulate peroxisomal abundance and suggest a yet unknown role of peroxisomes in flavivirus biology.
Keywords: Zika virus; capsid protein; dengue virus; flavivirus; human peroxin PEX19; peroxisomes.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





Similar articles
-
Flavivirus Infection Impairs Peroxisome Biogenesis and Early Antiviral Signaling.J Virol. 2015 Dec;89(24):12349-61. doi: 10.1128/JVI.01365-15. Epub 2015 Sep 30. J Virol. 2015. PMID: 26423946 Free PMC article.
-
Dengue and Zika virus NS4B proteins differ in topology and in determinants of ER membrane protein complex dependency.J Virol. 2025 Feb 25;99(2):e0144324. doi: 10.1128/jvi.01443-24. Epub 2024 Dec 31. J Virol. 2025. PMID: 39745435 Free PMC article.
-
Unlike dengue virus, the conserved 14-23 residues in N-terminal region of Zika virus capsid is not involved in lipid interactions.Biochim Biophys Acta Biomembr. 2020 Nov 1;1862(11):183440. doi: 10.1016/j.bbamem.2020.183440. Epub 2020 Aug 9. Biochim Biophys Acta Biomembr. 2020. PMID: 32783888
-
T Cell Immunity to Zika and Dengue Viral Infections.J Interferon Cytokine Res. 2017 Nov;37(11):475-479. doi: 10.1089/jir.2017.0106. J Interferon Cytokine Res. 2017. PMID: 29135369 Free PMC article. Review.
-
Immune Response to Dengue and Zika.Annu Rev Immunol. 2018 Apr 26;36:279-308. doi: 10.1146/annurev-immunol-042617-053142. Epub 2018 Jan 18. Annu Rev Immunol. 2018. PMID: 29345964 Free PMC article. Review.
Cited by
-
Peroxisomes and Viruses: Overview on Current Knowledge and Experimental Approaches.Methods Mol Biol. 2023;2643:271-294. doi: 10.1007/978-1-0716-3048-8_19. Methods Mol Biol. 2023. PMID: 36952192 Review.
-
Computational Insights on the Assembly of the Dengue Virus Membrane-Capsid-RNA Complex.J Membr Biol. 2025 Feb;258(1):75-96. doi: 10.1007/s00232-025-00337-4. Epub 2025 Jan 19. J Membr Biol. 2025. PMID: 39827433
-
Zika Virus Induces Degradation of the Numb Protein Required through Embryonic Neurogenesis.Viruses. 2023 May 27;15(6):1258. doi: 10.3390/v15061258. Viruses. 2023. PMID: 37376558 Free PMC article.
References
-
- Matsuzono Y., Kinoshita N., Tamura S., Shimozawa N., Hamasaki M., Ghaedi K., Wanders R.J.A., Suzuki Y., Kondo N., Fujiki Y. Human PEX19: cDNA cloning by functional complementation, mutation analysis in a patient with Zellweger syndrome, and potential role in peroxisomal membrane assembly. Proc. Natl. Acad. Sci. USA. 1999;96:2116–2121. doi: 10.1073/pnas.96.5.2116. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
