

Conservation and Divergence of SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) Gene Family between Wheat and Rice
- PMID: 35216210
- PMCID: PMC8874652
- DOI: 10.3390/ijms23042099
Conservation and Divergence of SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) Gene Family between Wheat and Rice
Abstract
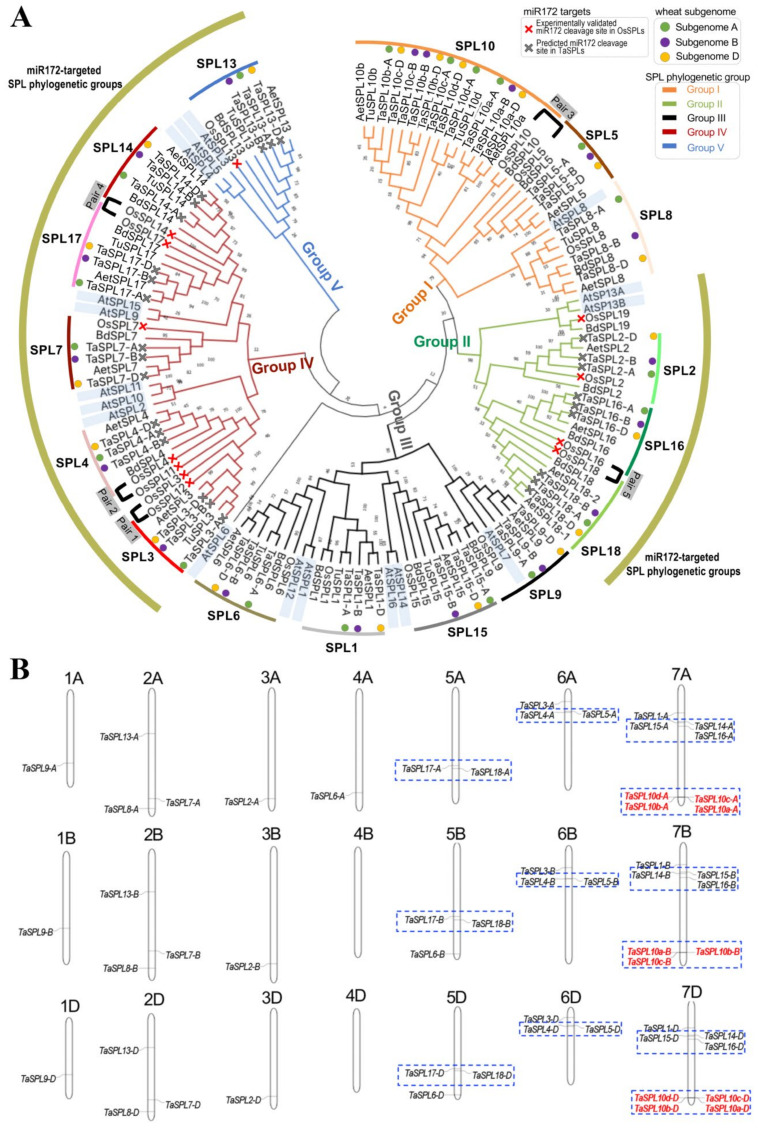
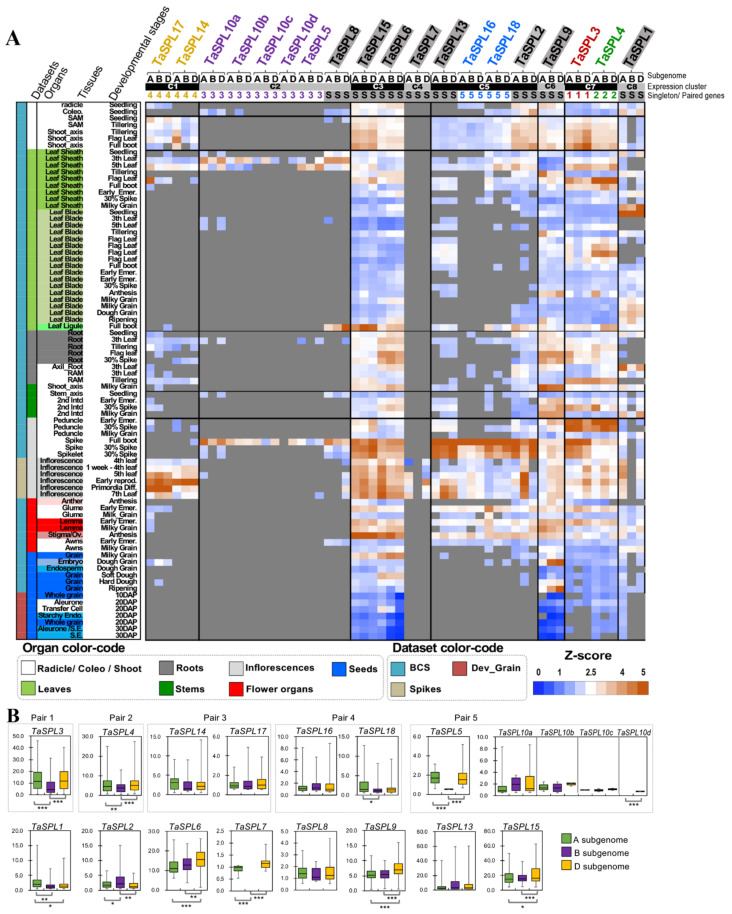
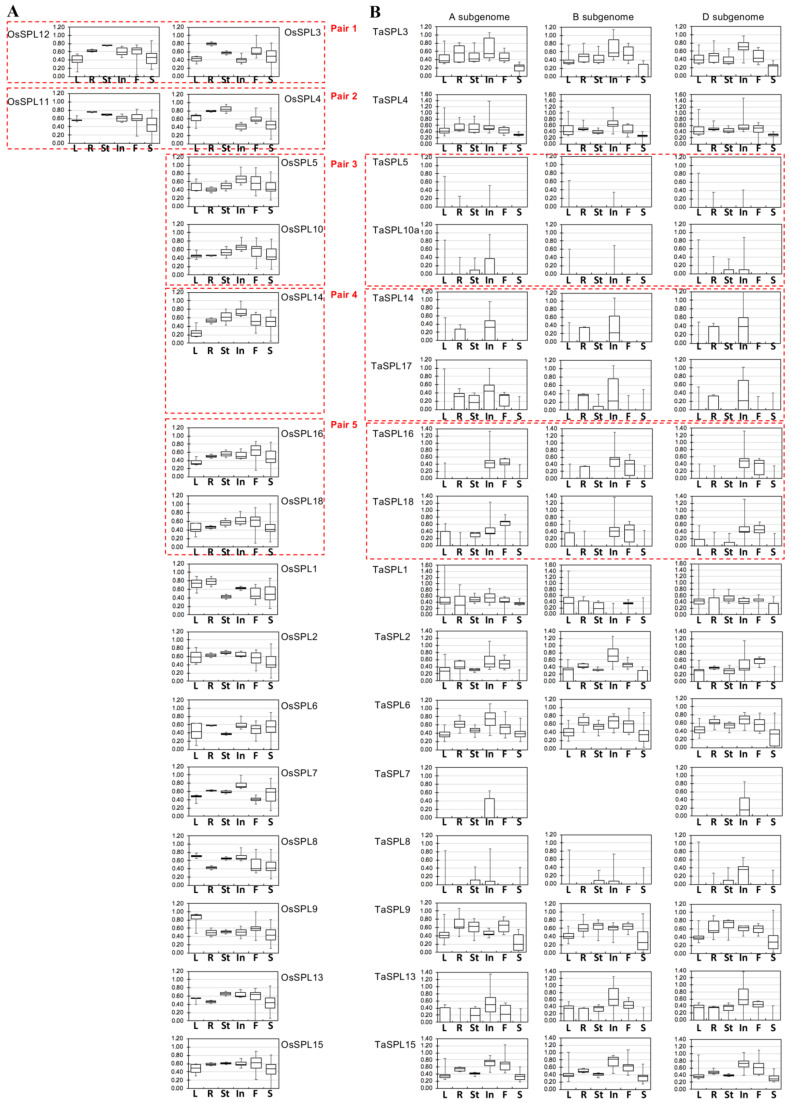
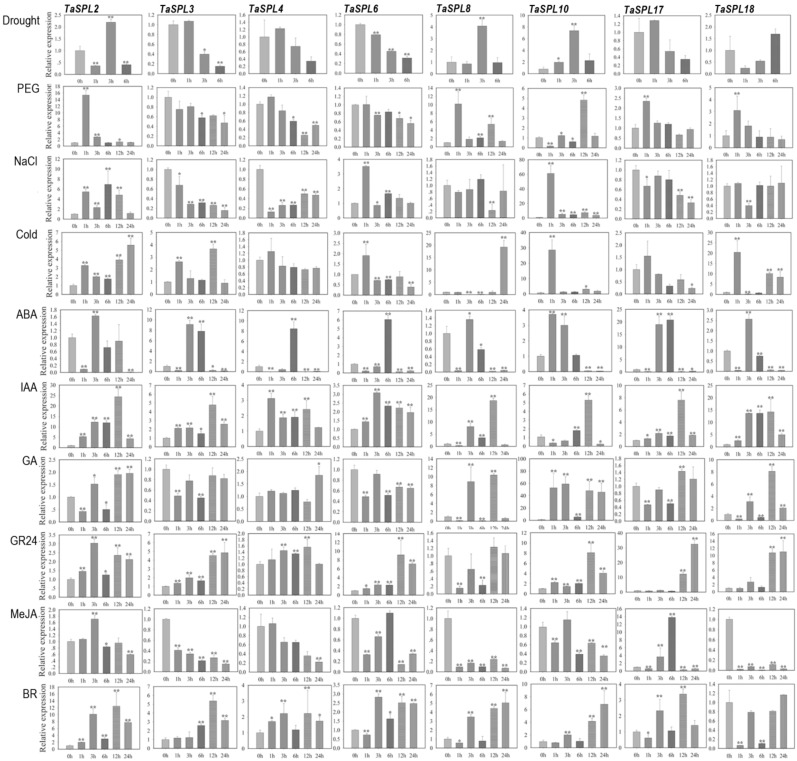
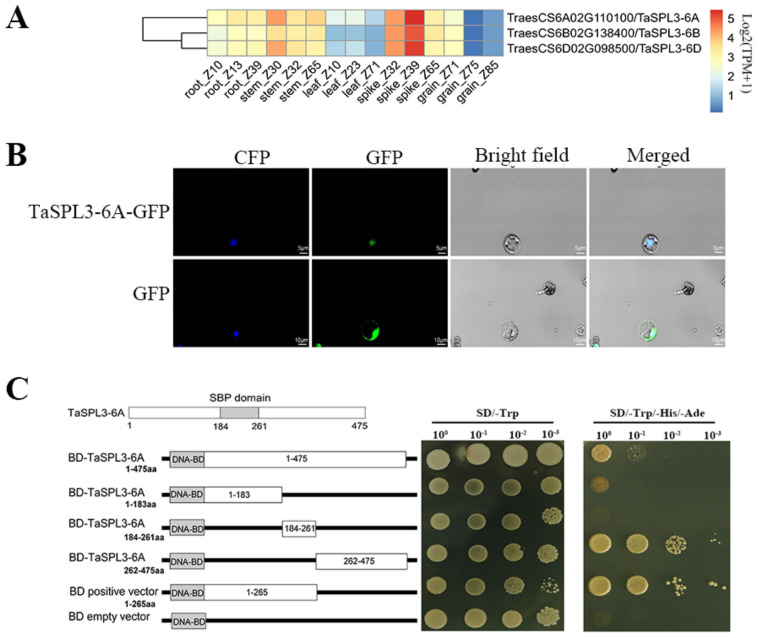
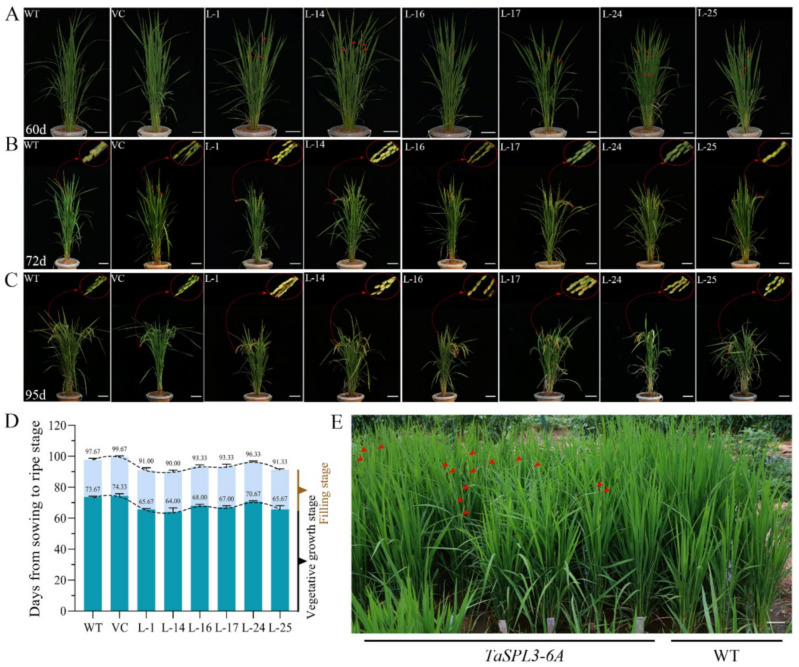
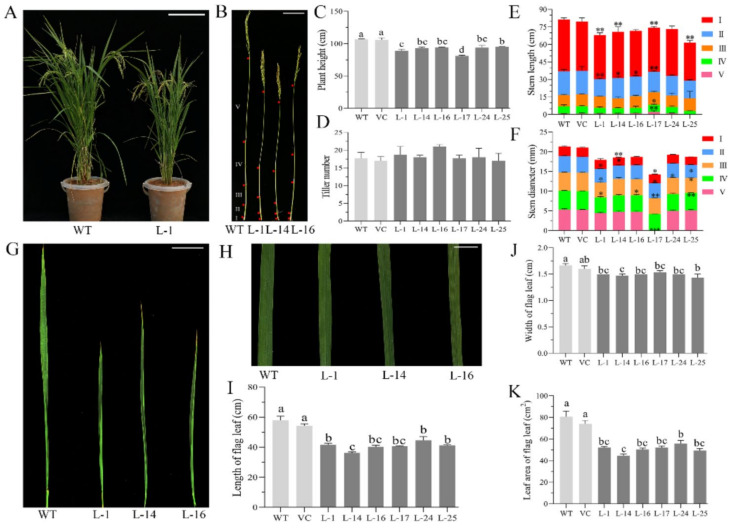
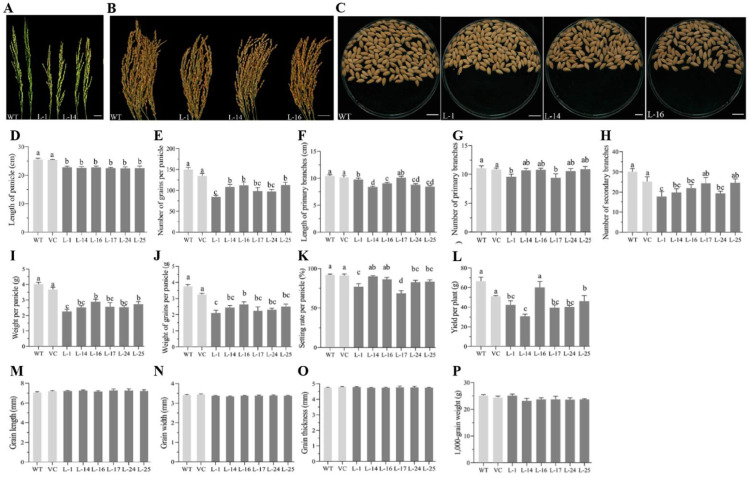
The SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) gene family affects plant architecture, panicle structure, and grain development, representing key genes for crop improvements. The objective of the present study is to utilize the well characterized SPLs' functions in rice to facilitate the functional genomics of TaSPL genes. To achieve these goals, we combined several approaches, including genome-wide analysis of TaSPLs, comparative genomic analysis, expression profiling, and functional study of TaSPL3 in rice. We established the orthologous relationships of 56 TaSPL genes with the corresponding OsSPLs, laying a foundation for the comparison of known SPL functions between wheat and rice. Some TaSPLs exhibited different spatial-temporal expression patterns when compared to their rice orthologs, thus implicating functional divergence. TaSPL2/6/8/10 were identified to respond to different abiotic stresses through the combination of RNA-seq and qPCR expression analysis. Additionally, ectopic expression of TaSPL3 in rice promotes heading dates, affects leaf and stem development, and leads to smaller panicles and decreased yields per panicle. In conclusion, our work provides useful information toward cataloging of the functions of TaSPLs, emphasized the conservation and divergence between TaSPLs and OsSPLs, and identified the important SPL genes for wheat improvement.
Keywords: Oryza sativa; SQUAMOSA-PROMOTER BINDING PROTEIN-LIKE (SPL) gene; Triticum aestivum; conservation and divergence; expression patterns; genome-wide analysis.
Conflict of interest statement
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manu-script, or in the decision to publish the results.
Figures
References
MeSH terms
Substances
Grants and funding
- 2016ZX08010004-004/National Genetically Modified New Varieties of Major Projects of China
- 31771418/National Natural Science Foundation of China
- 31371614/National Natural Science Foundation of China
- 2016CFB549/National Natural Science Foundation of Hubei China
- 2021XXJS070/Fundamental Research Funds for Central Universities, HUST
LinkOut - more resources
Full Text Sources
