

Tfh-cell-derived interleukin 21 sustains effector CD8+ T cell responses during chronic viral infection
- PMID: 35216666
- PMCID: PMC8916994
- DOI: 10.1016/j.immuni.2022.01.018
Tfh-cell-derived interleukin 21 sustains effector CD8+ T cell responses during chronic viral infection
Abstract
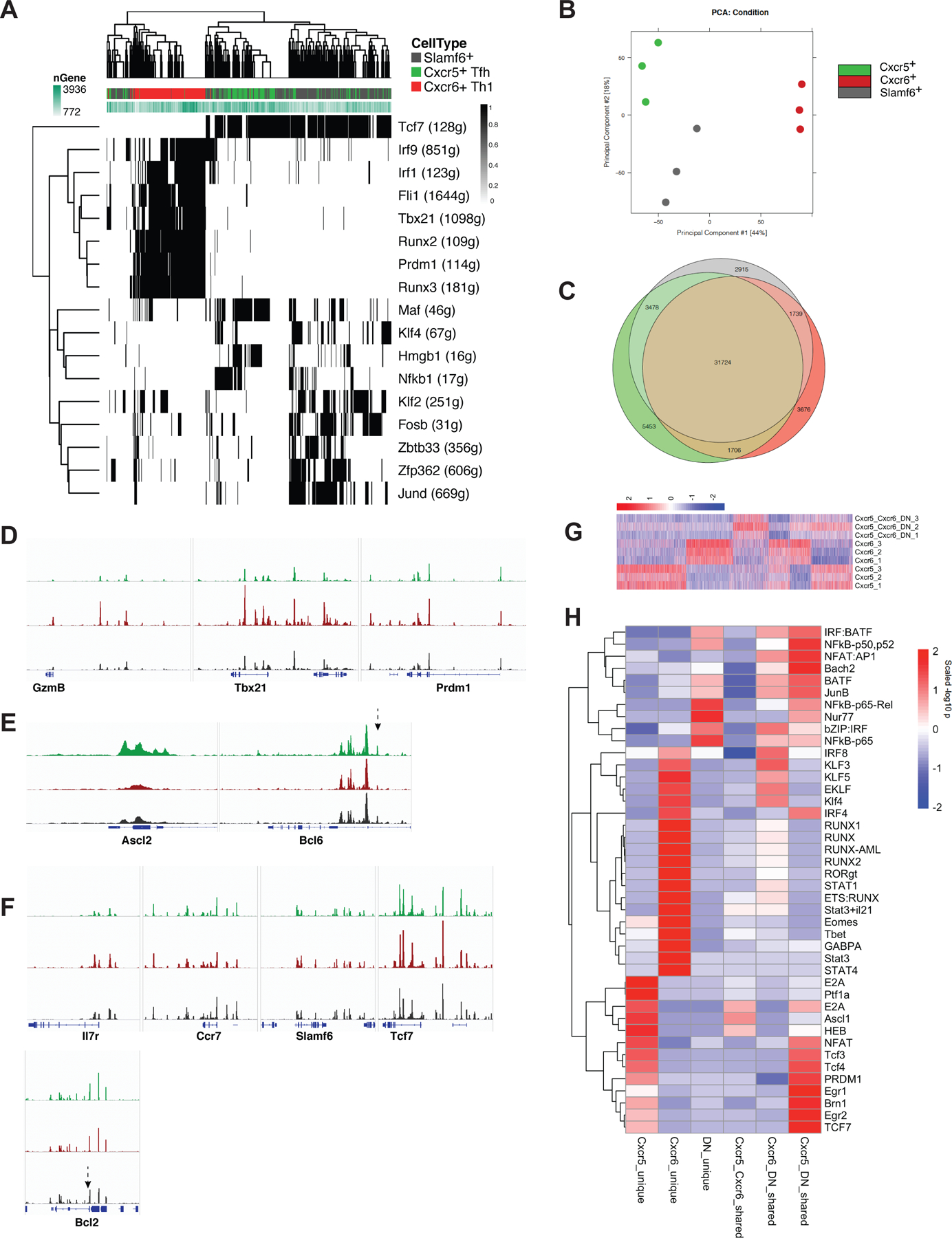
CD4+ T cell-derived interleukin 21 (IL-21) sustains CD8+ T cell responses during chronic viral infection, but the helper subset that confers this protection remains unclear. Here, we applied scRNA and ATAC-seq approaches to determine the heterogeneity of IL-21+CD4+ T cells during LCMV clone 13 infection. CD4+ T cells were comprised of three transcriptionally and epigenetically distinct populations: Cxcr6+ Th1 cells, Cxcr5+ Tfh cells, and a previously unrecognized Slamf6+ memory-like (Tml) subset. T cell differentiation was specifically redirected toward the Tml subset during chronic, but not acute, LCMV infection. Although this subset displayed an enhanced capacity to accumulate and some developmental plasticity, it remained largely quiescent, which may hinder its helper potential. Conversely, mixed bone marrow chimera experiments revealed that Tfh cell-derived IL-21 was critical to sustain CD8+ T cell responses and viral control. Thus, strategies that bolster IL-21+Tfh cell responses may prove effective in enhancing CD8+ T cell-mediated immunity.
Keywords: ATAC-seq; CD4 T cells; IL-21; LCMV Cl13; single-cell RNA sequencing.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures






References
Publication types
MeSH terms
Substances
Grants and funding
- R01 AI152443/AI/NIAID NIH HHS/United States
- UL1 TR001863/TR/NCATS NIH HHS/United States
- F30 DK127526/DK/NIDDK NIH HHS/United States
- F30 CA246920/CA/NCI NIH HHS/United States
- R00 AI153537/AI/NIAID NIH HHS/United States
- R01 AI148403/AI/NIAID NIH HHS/United States
- P30 DK045735/DK/NIDDK NIH HHS/United States
- T32 GM080202/GM/NIGMS NIH HHS/United States
- K99 AI153537/AI/NIAID NIH HHS/United States
- R01 AI156290/AI/NIAID NIH HHS/United States
- R01 AI125741/AI/NIAID NIH HHS/United States
- R37 AR040072/AR/NIAMS NIH HHS/United States
- R01 AR074545/AR/NIAMS NIH HHS/United States
- R01 AI049360/AI/NIAID NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
