

PRDM9 losses in vertebrates are coupled to those of paralogs ZCWPW1 and ZCWPW2
- PMID: 35217607
- PMCID: PMC8892340
- DOI: 10.1073/pnas.2114401119
PRDM9 losses in vertebrates are coupled to those of paralogs ZCWPW1 and ZCWPW2
Abstract
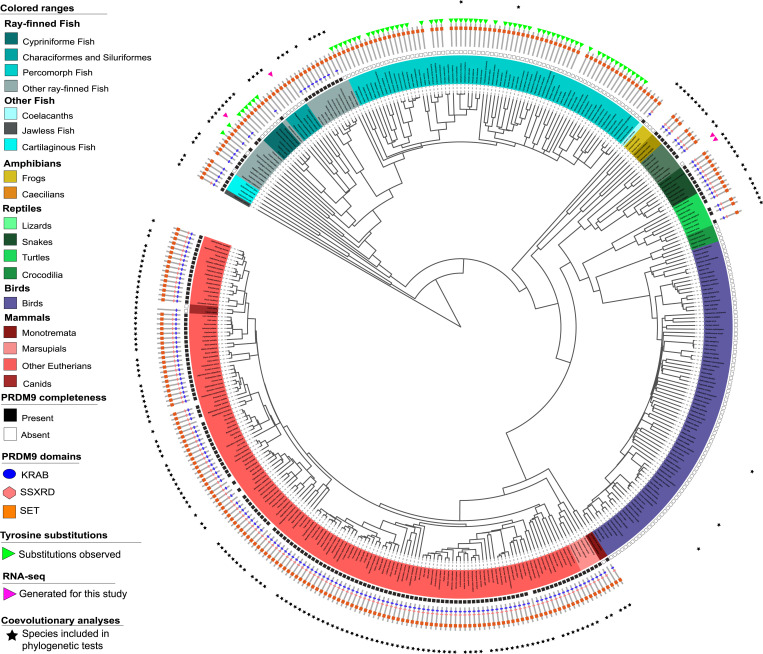
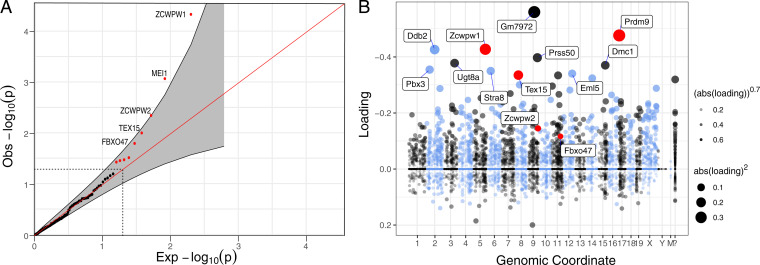
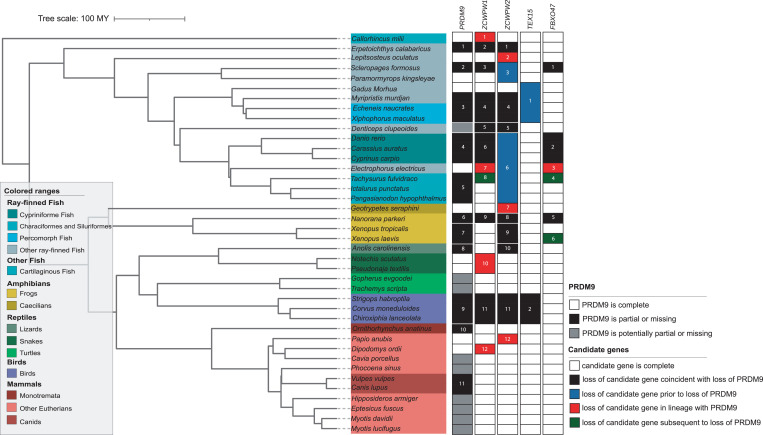
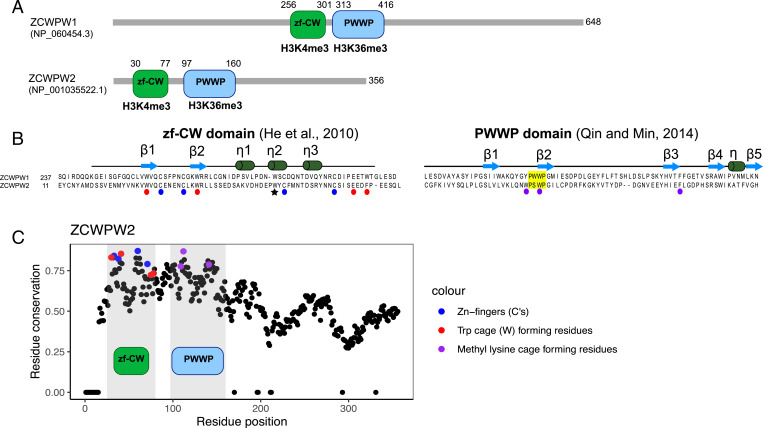
In most mammals and likely throughout vertebrates, the gene PRDM9 specifies the locations of meiotic double strand breaks; in mice and humans at least, it also aids in their repair. For both roles, many of the molecular partners remain unknown. Here, we take a phylogenetic approach to identify genes that may be interacting with PRDM9 by leveraging the fact that PRDM9 arose before the origin of vertebrates but was lost many times, either partially or entirely-and with it, its role in recombination. As a first step, we characterize PRDM9 domain composition across 446 vertebrate species, inferring at least 13 independent losses. We then use the interdigitation of PRDM9 orthologs across vertebrates to test whether it coevolved with any of 241 candidate genes coexpressed with PRDM9 in mice or associated with recombination phenotypes in mammals. Accounting for the phylogenetic relationship among a subsample of 189 species, we find two genes whose presence and absence is unexpectedly coincident with that of PRDM9: ZCWPW1, which was recently shown to facilitate double strand break repair, and its paralog ZCWPW2, as well as, more tentatively, TEX15 and FBXO47ZCWPW2 is expected to be recruited to sites of PRDM9 binding; its tight coevolution with PRDM9 across vertebrates suggests that it is a key interactor within mammals and beyond, with a role either in recruiting the recombination machinery or in double strand break repair.
Keywords: PRDM9 evolution; comparative genomics; genetics; phylogenetics; recombination.
Copyright © 2022 the Author(s). Published by PNAS.
Conflict of interest statement
The authors declare no competing interest.
Figures
References
-
- Keeney S., Neale M. J., Initiation of meiotic recombination by formation of DNA double-strand breaks: Mechanism and regulation. Biochem. Soc. Trans. 34, 523–525 (2006). - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
