

Causal Evidence for a Role of Cerebellar Lobulus Simplex in Prefrontal-Hippocampal Interaction in Spatial Working Memory Decision-Making
- PMID: 35218525
- PMCID: PMC10230449
- DOI: 10.1007/s12311-022-01383-7
Causal Evidence for a Role of Cerebellar Lobulus Simplex in Prefrontal-Hippocampal Interaction in Spatial Working Memory Decision-Making
Abstract
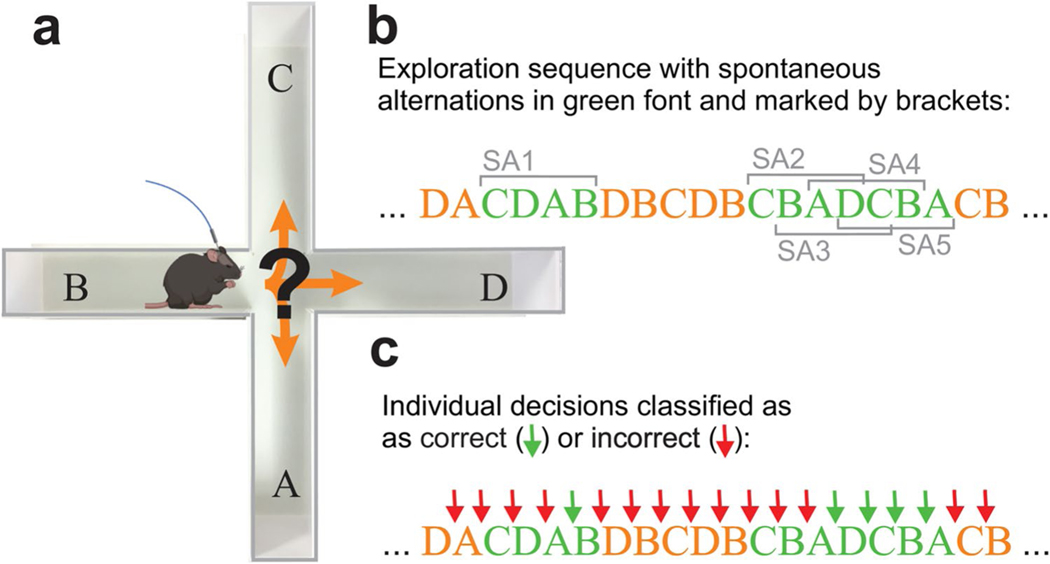
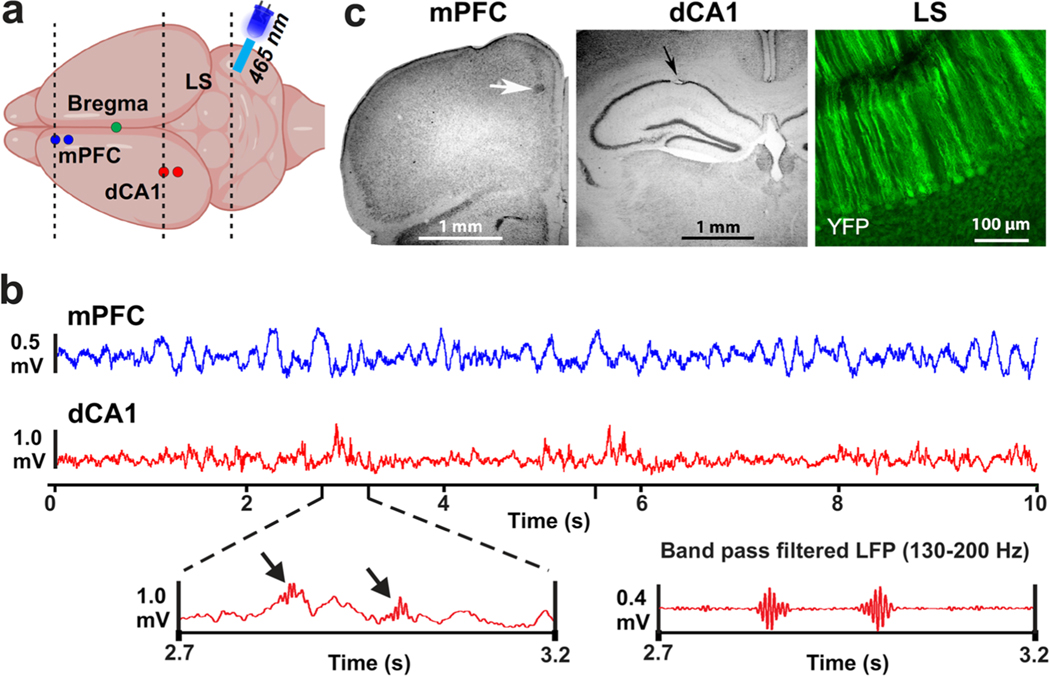
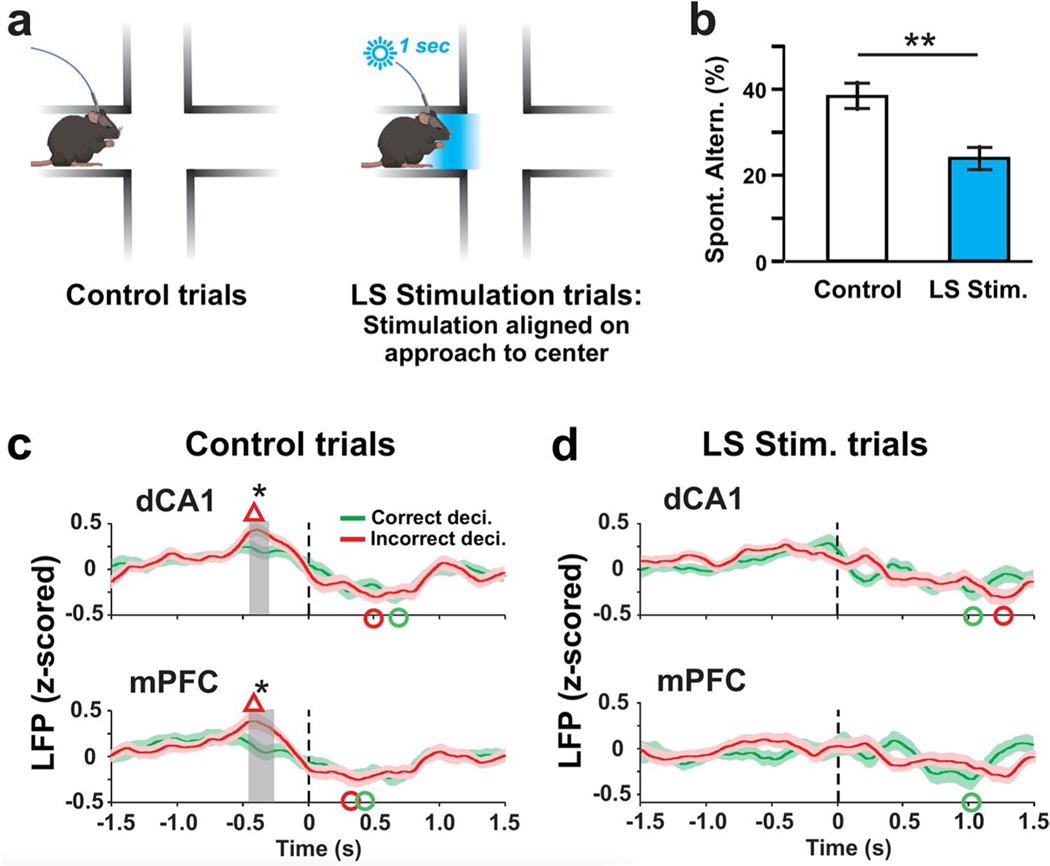
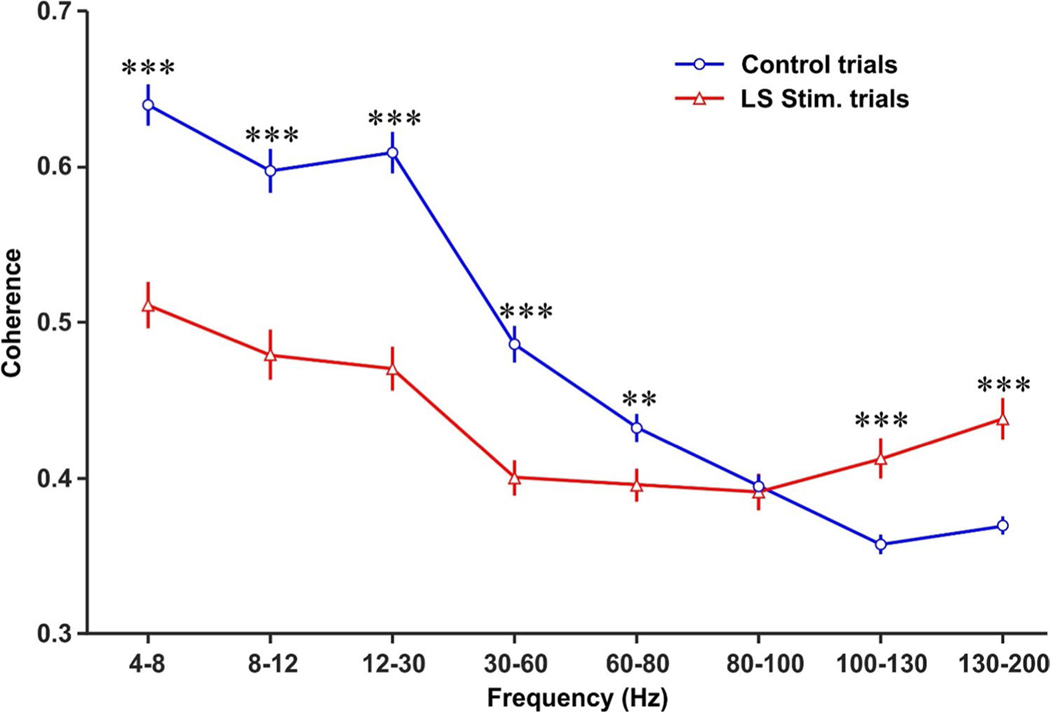
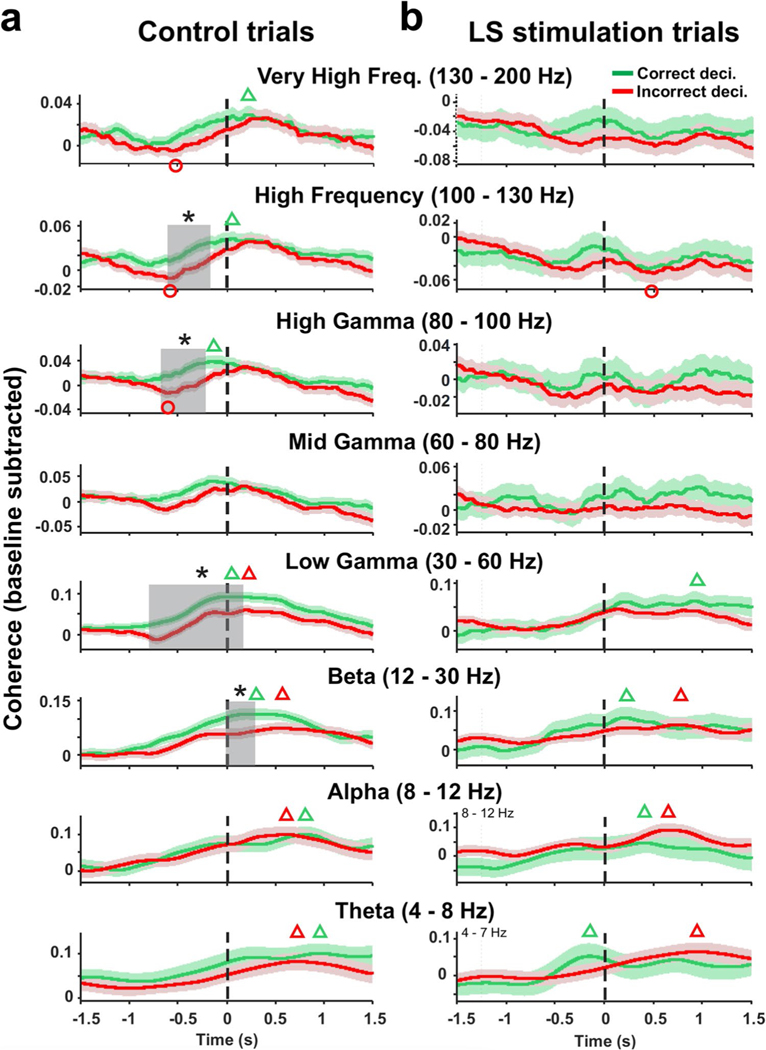
Spatial working memory (SWM) is a cerebrocerebellar cognitive skill supporting survival-relevant behaviors, such as optimizing foraging behavior by remembering recent routes and visited sites. It is known that SWM decision-making in rodents requires the medial prefrontal cortex (mPFC) and dorsal hippocampus. The decision process in SWM tasks carries a specific electrophysiological signature of a brief, decision-related increase in neuronal communication in the form of an increase in the coherence of neuronal theta oscillations (4-12 Hz) between the mPFC and dorsal hippocampus, a finding we replicated here during spontaneous exploration of a plus maze in freely moving mice. We further evaluated SWM decision-related coherence changes within frequency bands above theta. Decision-related coherence increases occurred in seven frequency bands between 4 and 200 Hz and decision-outcome-related differences in coherence modulation occurred within the beta and gamma frequency bands and in higher frequency oscillations up to 130 Hz. With recent evidence that Purkinje cells in the cerebellar lobulus simplex (LS) represent information about the phase and phase differences of gamma oscillations in the mPFC and dorsal hippocampus, we hypothesized that LS might be involved in the modulation of mPFC-hippocampal gamma coherence. We show that optical stimulation of LS significantly impairs SWM performance and decision-related mPFC-dCA1 coherence modulation, providing causal evidence for an involvement of cerebellar LS in SWM decision-making at the behavioral and neuronal level. Our findings suggest that the cerebellum might contribute to SWM decision-making by optimizing the decision-related modulation of mPFC-dCA1 coherence.
Keywords: Cerebellum; Cognition; Coherence; Decision-making; Electrophysiology; Hippocampus; Medial prefrontal cortex; Neuronal communication; Optogenetics; Prefrontal cortex; Purkinje cell; Spatial working memory.
© 2022. The Author(s), under exclusive licence to Springer Science+Business Media, LLC, part of Springer Nature.
Conflict of interest statement
Declarations
Figures
Similar articles
-
Ventral Midline Thalamus Is Critical for Hippocampal-Prefrontal Synchrony and Spatial Working Memory.J Neurosci. 2016 Aug 10;36(32):8372-89. doi: 10.1523/JNEUROSCI.0991-16.2016. J Neurosci. 2016. PMID: 27511010 Free PMC article.
-
Adaptive closed-loop modulation of cortical theta oscillations: Insights into the neural dynamics of navigational decision-making.Brain Stimul. 2024 Sep-Oct;17(5):1101-1118. doi: 10.1016/j.brs.2024.09.005. Epub 2024 Sep 12. Brain Stimul. 2024. PMID: 39277130
-
Theta oscillations in the medial prefrontal cortex are modulated by spatial working memory and synchronize with the hippocampus through its ventral subregion.J Neurosci. 2013 Aug 28;33(35):14211-24. doi: 10.1523/JNEUROSCI.2378-13.2013. J Neurosci. 2013. PMID: 23986255 Free PMC article.
-
Role of the thalamic nucleus reuniens in mediating interactions between the hippocampus and medial prefrontal cortex during spatial working memory.Front Syst Neurosci. 2015 Mar 10;9:29. doi: 10.3389/fnsys.2015.00029. eCollection 2015. Front Syst Neurosci. 2015. PMID: 25805977 Free PMC article. Review.
-
The nucleus reuniens orchestrates prefrontal-hippocampal synchrony during spatial working memory.Neurosci Biobehav Rev. 2021 Sep;128:415-420. doi: 10.1016/j.neubiorev.2021.05.033. Epub 2021 Jul 1. Neurosci Biobehav Rev. 2021. PMID: 34217746 Free PMC article. Review.
Cited by
-
Cerebellar control of thalamocortical circuits for cognitive function: A review of pathways and a proposed mechanism.Front Syst Neurosci. 2023 Mar 30;17:1126508. doi: 10.3389/fnsys.2023.1126508. eCollection 2023. Front Syst Neurosci. 2023. PMID: 37064161 Free PMC article. Review.
-
Cerebellum-Cortical Interaction in Spatial Navigation and Its Alteration in Dementias.Brain Sci. 2022 Apr 20;12(5):523. doi: 10.3390/brainsci12050523. Brain Sci. 2022. PMID: 35624910 Free PMC article. Review.
-
Serotonin Modulation of Dorsoventral Hippocampus in Physiology and Schizophrenia.Int J Mol Sci. 2025 Jul 27;26(15):7253. doi: 10.3390/ijms26157253. Int J Mol Sci. 2025. PMID: 40806385 Free PMC article. Review.
-
Task-specific oscillatory synchronization of prefrontal cortex, nucleus reuniens, and hippocampus during working memory.iScience. 2023 Aug 3;26(9):107532. doi: 10.1016/j.isci.2023.107532. eCollection 2023 Sep 15. iScience. 2023. PMID: 37636046 Free PMC article.
-
Social memory deficit caused by dysregulation of the cerebellar vermis.Nat Commun. 2023 Sep 26;14(1):6007. doi: 10.1038/s41467-023-41744-2. Nat Commun. 2023. PMID: 37752149 Free PMC article.
References
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
