

Dendritic Spine Density Scales with Microtubule Number in Rat Hippocampal Dendrites
- PMID: 35218884
- PMCID: PMC9038701
- DOI: 10.1016/j.neuroscience.2022.02.021
Dendritic Spine Density Scales with Microtubule Number in Rat Hippocampal Dendrites
Abstract
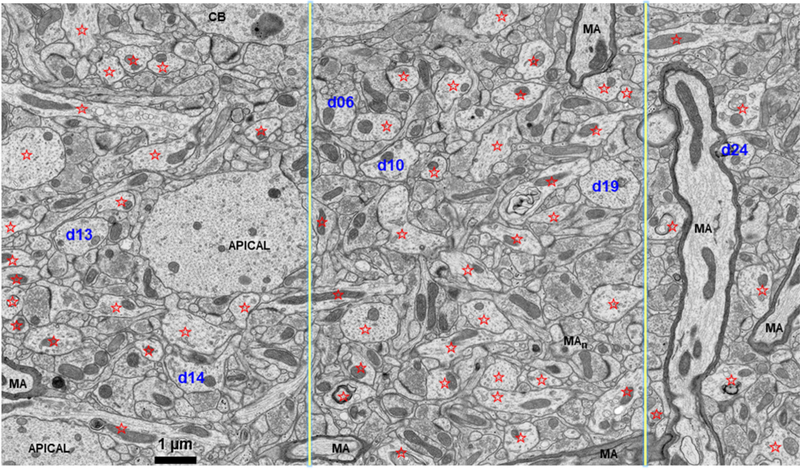
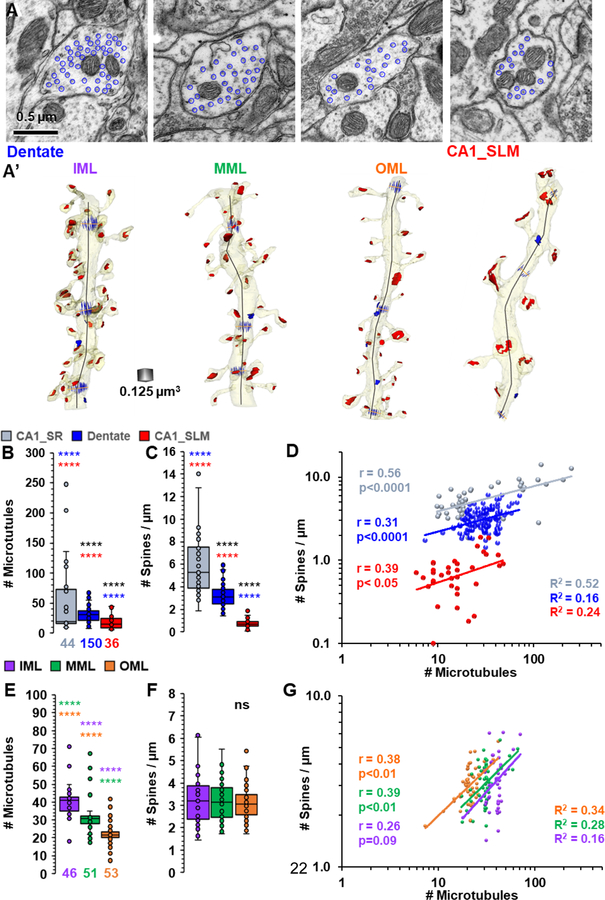
Microtubules deliver essential resources to and from synapses. Three-dimensional reconstructions in rat hippocampus reveal a sampling bias regarding spine density that needs to be controlled for dendrite caliber and resource delivery based on microtubule number. The strength of this relationship varies across dendritic arbors, as illustrated for area CA1 and dentate gyrus. In both regions, proximal dendrites had more microtubules than distal dendrites. For CA1 pyramidal cells, spine density was greater on thicker than thinner dendrites in stratum radiatum, or on the more uniformly thin terminal dendrites in stratum lacunosum moleculare. In contrast, spine density was constant across the cone shaped arbor of tapering dendrites from dentate granule cells. These differences suggest that thicker dendrites supply microtubules to subsequent dendritic branches and local dendritic spines, whereas microtubules in thinner dendrites need only provide resources to local spines. Most microtubules ran parallel to dendrite length and associated with long, presumably stable mitochondria, which occasionally branched into lateral dendritic branches. Short, presumably mobile, mitochondria were tethered to microtubules that bent and appeared to direct them into a thin lateral branch. Prior work showed that dendritic segments with the same number of microtubules had elevated resources in subregions of their dendritic shafts where spine synapses had enlarged, and spine clusters had formed. Thus, additional microtubules were not required for redistribution of resources locally to growing spines or synapses. These results provide new understanding about the potential for microtubules to regulate resource delivery to and from dendritic branches and locally among dendritic spines.
Keywords: 3D-reconstruction; microtubules; spine density; synapses; ultrastructure.
Copyright © 2022 IBRO. Published by Elsevier Ltd. All rights reserved.
Figures



References
-
- Ehlers MD, Dendritic trafficking for neuronal growth and plasticity, Biochem. Soc. Trans, 41 (2013) 1365–1382. - PubMed
-
- Fiala JC, Kirov SA, Feinberg MD, Petrak LJ, George P, Goddard CA, Harris KM, Timing of neuronal and glial ultrastructure disruption during brain slice preparation and recovery in vitro, J Comp Neurol, 465 (2003) 90–103. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
