

Mucin 5AC-Mediated CD44/ITGB1 Clustering Mobilizes Adipose-Derived Mesenchymal Stem Cells to Modulate Pancreatic Cancer Stromal Heterogeneity
- PMID: 35219699
- PMCID: PMC9117481
- DOI: 10.1053/j.gastro.2022.02.032
Mucin 5AC-Mediated CD44/ITGB1 Clustering Mobilizes Adipose-Derived Mesenchymal Stem Cells to Modulate Pancreatic Cancer Stromal Heterogeneity
Abstract
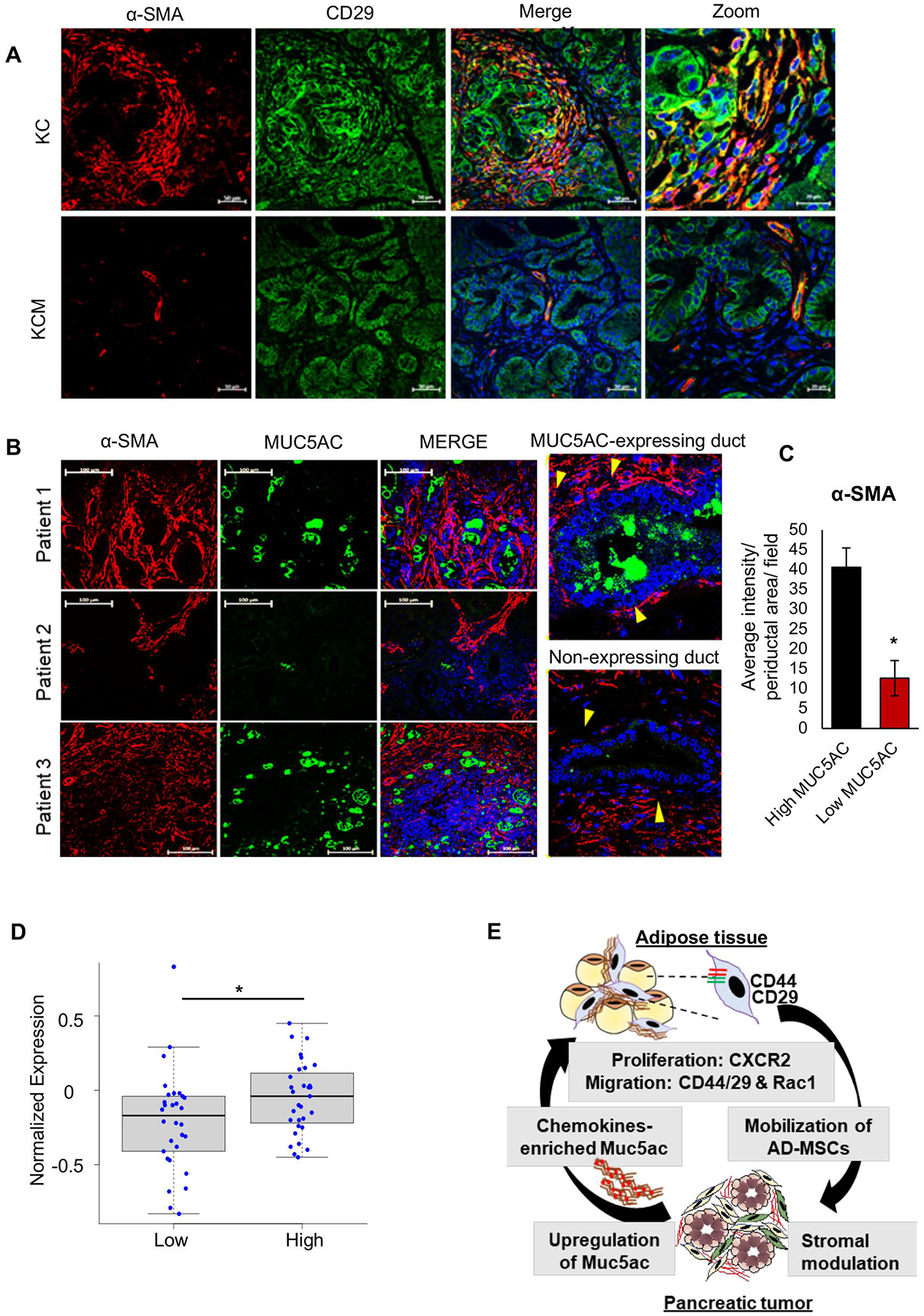
Background & aims: Secreted mucin 5AC (MUC5AC) promotes pancreatic cancer (PC) progression and chemoresistance, suggesting its clinical association with poor prognosis. RNA sequencing analysis from the autochthonous pancreatic tumors showed a significant stromal alteration on genetic ablation of Muc5ac. Previously, depletion or targeting the stromal fibroblasts showed an ambiguous effect on PC pathogenesis. Hence, identifying the molecular players and mechanisms driving fibroblast heterogeneity is critical for improved clinical outcomes.
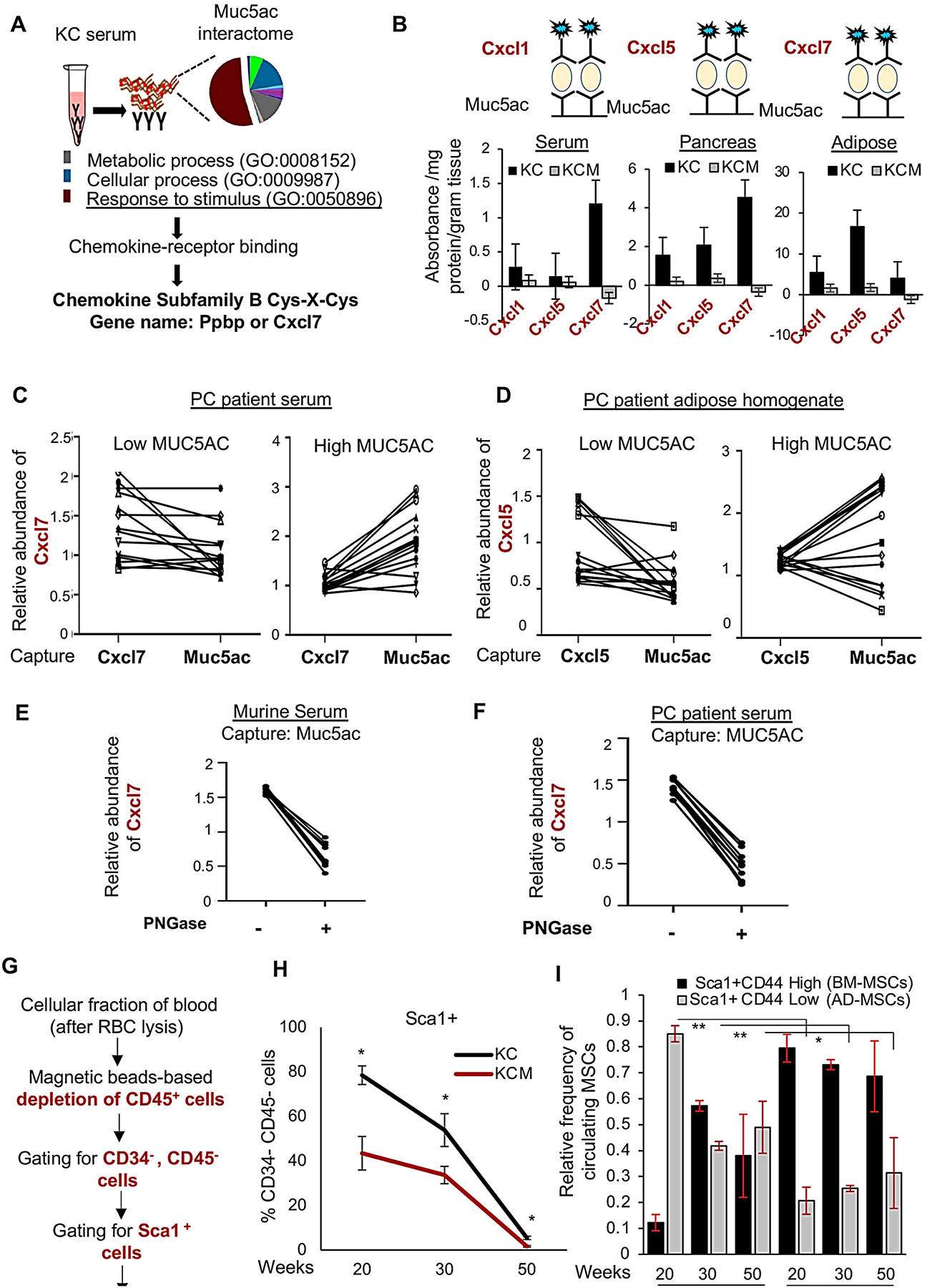
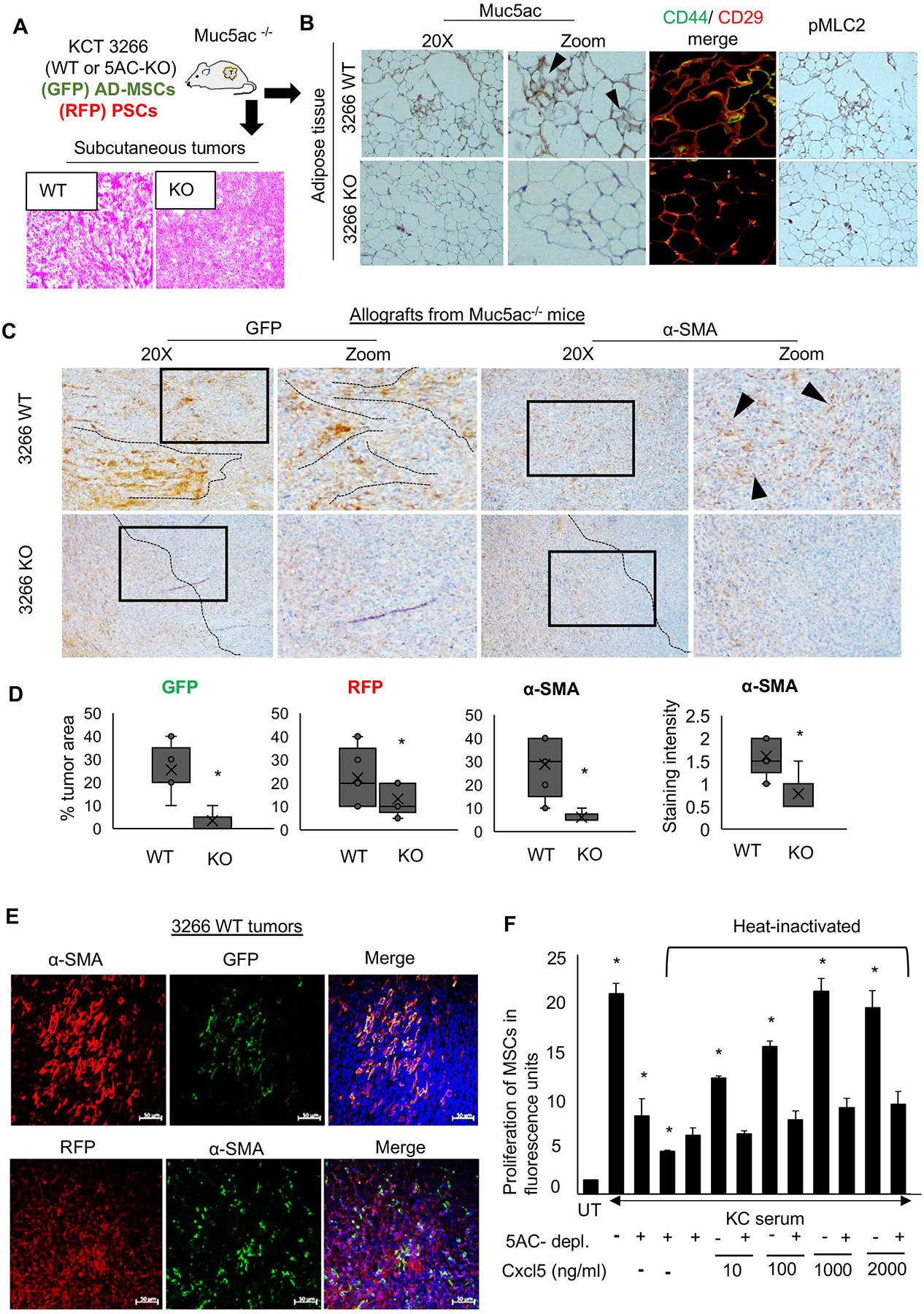
Methods: Autochthonous murine models of PC (KrasG12D, Pdx1-Cre [KC] and KrasG12D, Pdx1-Cre, Muc5ac-/- [KCM]) and co-implanted allografts of murine PC cell lines (Muc5ac wild-type and CRISPR/Cas knockout) with adipose-derived mesenchymal stem cells (AD-MSCs) were used to assess the role of Muc5ac in stromal heterogeneity. Proliferation, migration, and surface expression of cell-adhesion markers on AD-MSCs were measured using live-cell imaging and flow cytometry. MUC5AC-interactome was investigated using mass-spectrometry and enzyme-linked immunosorbent assay.
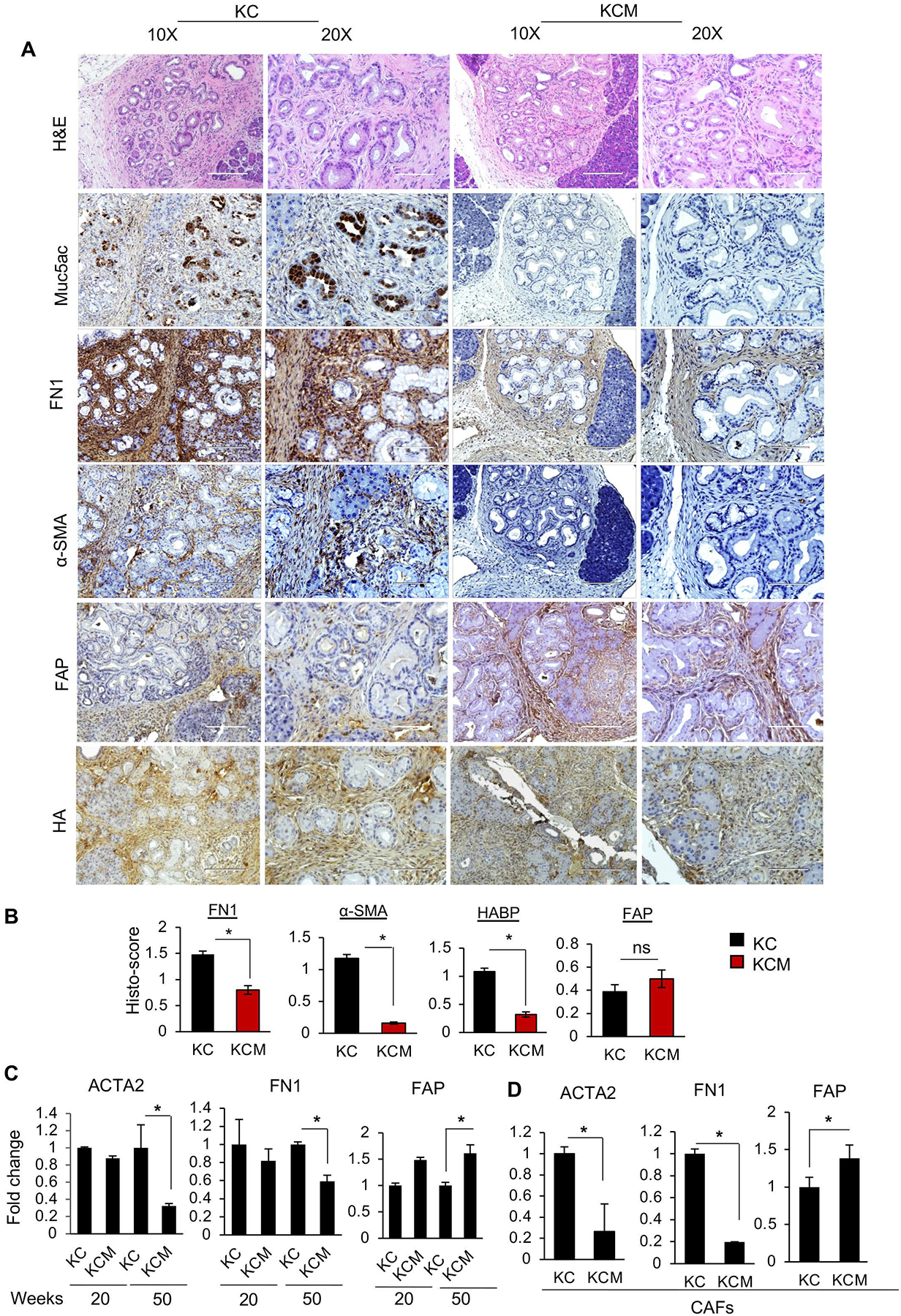
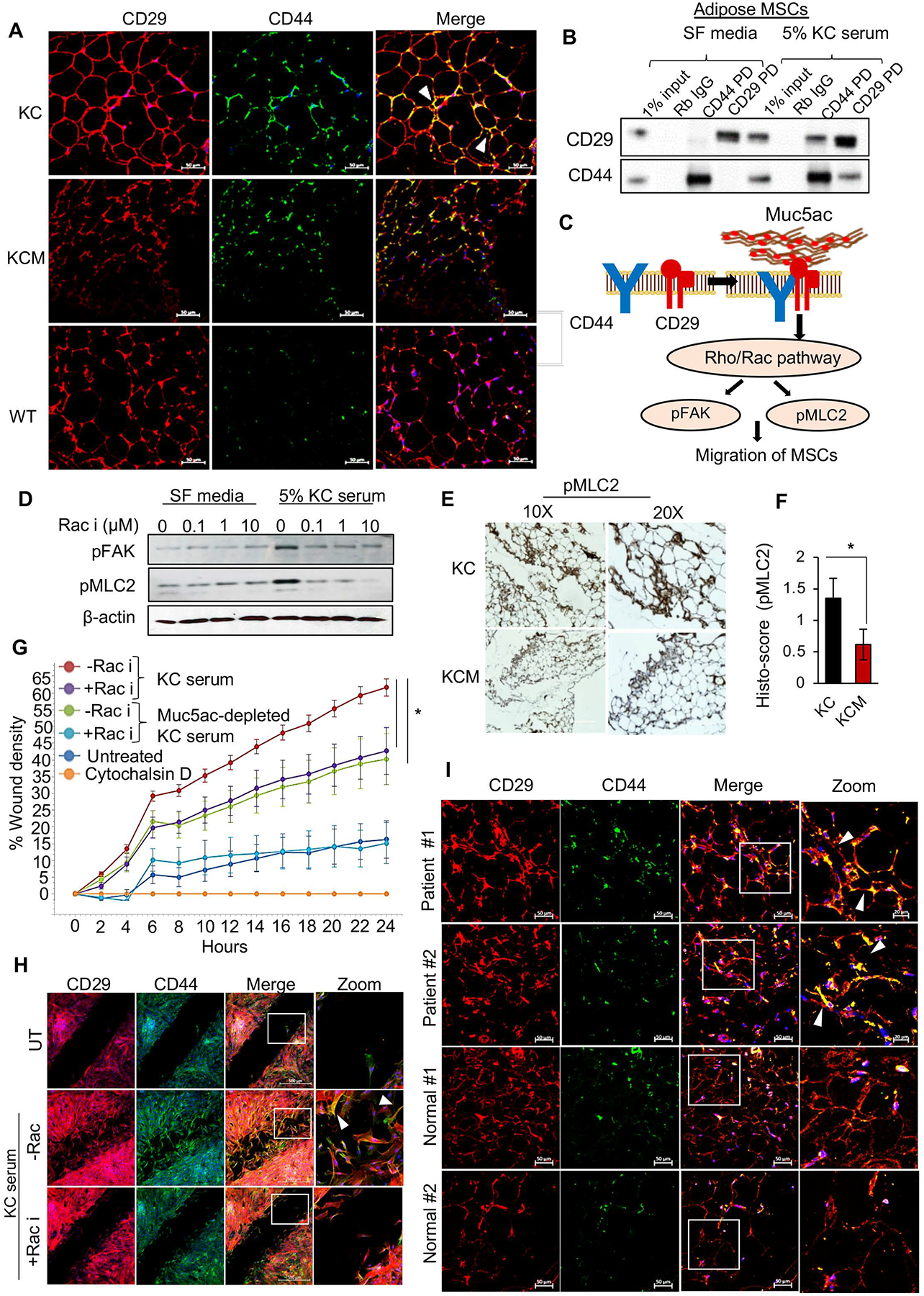
Results: The KCM tumors showed a significant decrease in the expression of α-smooth muscle actin and fibronectin compared with histology-matched KC tumors. Our study showed that MUC5AC, carrying tumor secretome, gets enriched in the adipose tissues of tumor-bearing mice and patients with PC, promoting CD44/CD29 (integrin-β1) clustering that leads to Rac1 activation and migration of AD-MSCs. Furthermore, treatment with KC-derived serum enhanced proliferation and migration of AD-MSCs, which was abolished on Muc5ac-depletion or pharmacologic inhibition of CXCR2 and Rac1, respectively. The AD-MSCs significantly contribute toward α-smooth muscle actin-positive cancer-associated fibroblasts population in Muc5ac-dependent manner, as suggested by autochthonous tumors, co-implantation xenografts, and patient tumors.
Conclusion: MUC5AC, secreted during PC progression, enriches in adipose and enhances the mobilization of AD-MSCs. On recruitment to pancreatic tumors, AD-MSCs proliferate and contribute towards stromal heterogeneity.
Keywords: Cancer-Associated Fibroblast; Chemokines; MUC5AC; Mesenchymal Stem Cells; Pancreatic Cancer; Stroma.
Copyright © 2022 AGA Institute. Published by Elsevier Inc. All rights reserved.
Conflict of interest statement
Figures

References
-
- Neoptolemos JP, Kleeff J, Michl P, et al. Therapeutic developments in pancreatic cancer: current and future perspectives. Nature reviews Gastroenterology & hepatology 2018;15:333–348. - PubMed
-
- Neesse A, Bauer CA, Öhlund D, et al. Stromal biology and therapy in pancreatic cancer: ready for clinical translation? Gut 2019;68:159–171. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous
