

Melatonin enhances osteoblastogenesis of senescent bone marrow stromal cells through NSD2-mediated chromatin remodelling
- PMID: 35220680
- PMCID: PMC8882236
- DOI: 10.1002/ctm2.746
Melatonin enhances osteoblastogenesis of senescent bone marrow stromal cells through NSD2-mediated chromatin remodelling
Abstract
Background: Aging-associated osteoporosis is frequently seen in the elderly in clinic, but efficient managements are limited because of unclear nosogenesis. The current study aims to investigate the role of melatonin on senescent bone marrow stromal cells (BMSCs) and the underlying regulating mechanism.
Methods: Melatonin levels were tested by ELISA. Gene expression profiles were performed by RNA-sequencing, enrichment of H3K36me2 on gene promoters was analyzed by Chromatin Immunoprecipitation Sequencing (ChIP-seq), and chromatin accessibility was determined by Assay for Transposase-Accessible Chromatin with high-throughput sequencing (ATAC-seq). Osteogenesis of BMSCs in vitro was measured by Alizarin Red and Alkaline Phosphatase staining, and in vivo effects of melatonin was assessed by histological staining and micro computed tomography (micro-CT) scan. Correlation of NSD2 expression and severity of senile osteoporosis patients were analyzed by Pearson correlation.
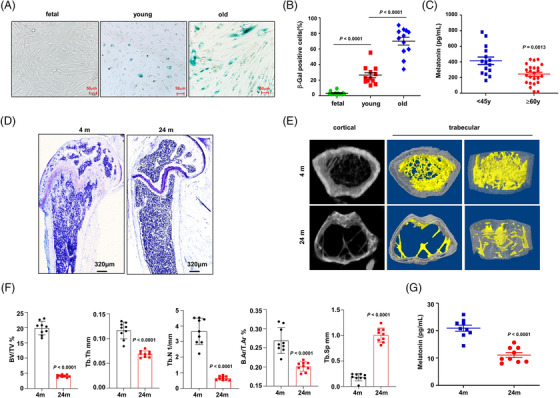
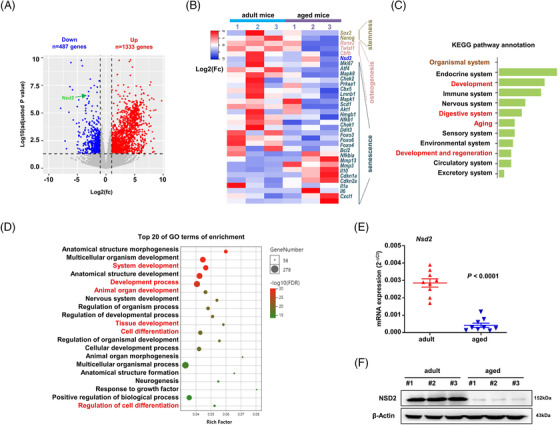
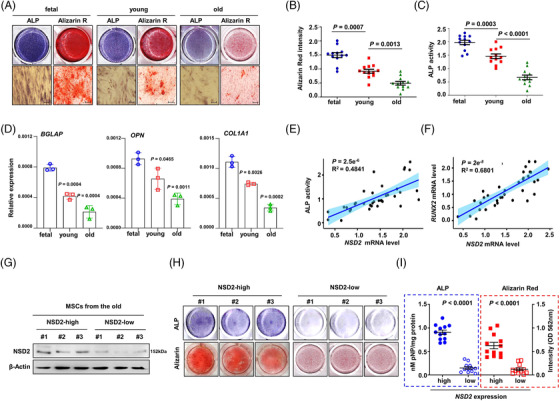
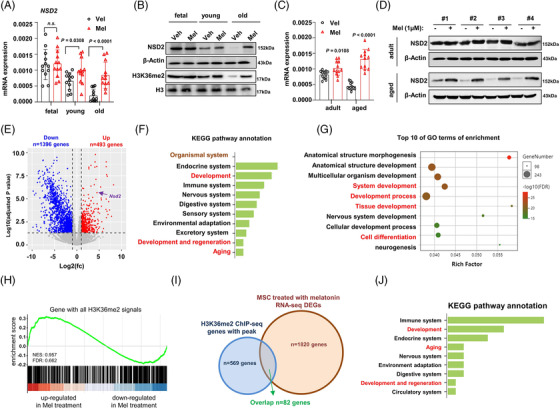
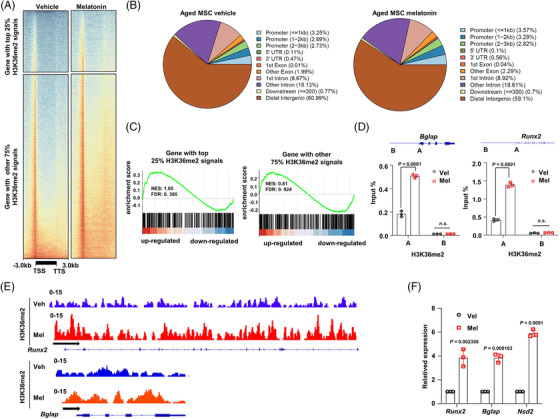
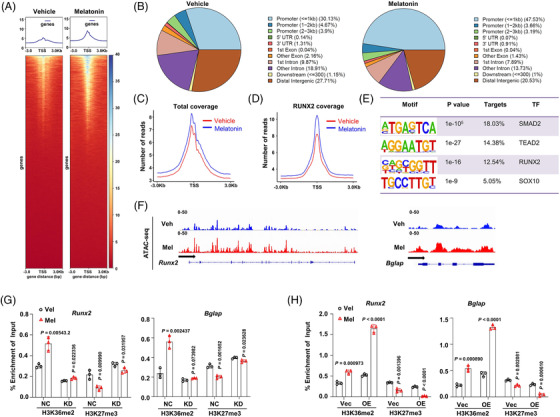
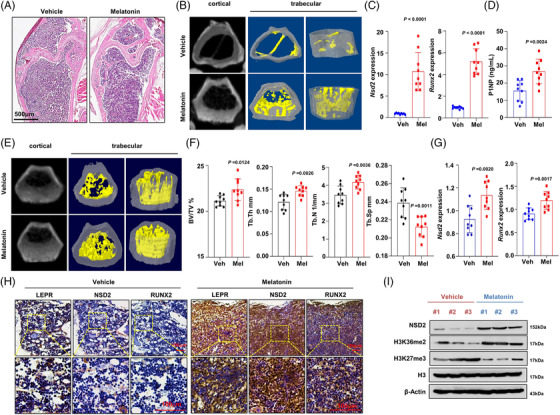
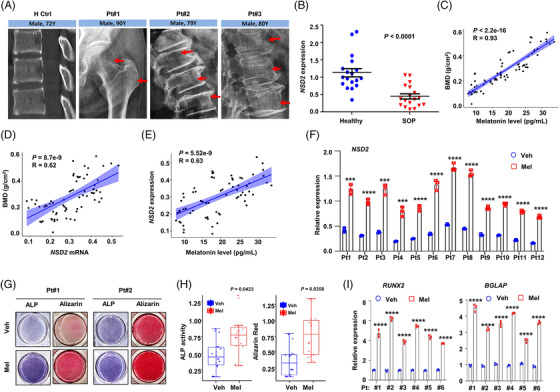
Results: Melatonin levels were decreased during aging in human bone marrow, accompanied by downregulation of the histone methyltransferase nuclear receptor binding SET domain protein 2 (NSD2) expression in the senescent BMSCs. Melatonin stimulated the expression of NSD2 through MT1/2-mediated signaling pathways, resulting in the rebalancing of H3K36me2 and H3K27me3 modifications to increase chromatin accessibility of the osteogenic genes, runt-related transcription factor 2 (RUNX2) and bone gamma-carboxyglutamate protein (BGLAP). Melatonin promoted osteogenesis of BMSCs in vitro, and alleviates osteoporosis progression in the aging mice. In clinic, severity of senile osteoporosis (SOP) was negatively correlated with melatonin level in bone marrow, as well as NSD2 expression in BMSCs. Similarly, melatonin remarkably enhanced osteogenic differentiation of BMSCs derived from SOP patients in vitro.
Conclusions: Collectively, our study dissects previously unreported mechanistic insights into the epigenetic regulating machinery of melatonin in meliorating osteogenic differentiation of senescent BMSC, and provides evidence for application of melatonin in preventing aging-associated bone loss.
Keywords: NSD2; bone marrow stromal cells; melatonin; osteoporosis; senescence.
© 2022 The Authors. Clinical and Translational Medicine published by John Wiley & Sons Australia, Ltd on behalf of Shanghai Institute of Clinical Bioinformatics.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
Similar articles
-
Rejuvenation of Bone Marrow Mesenchymal Stem Cells: Mechanisms and Their Application in Senile Osteoporosis Treatment.Biomolecules. 2025 Feb 13;15(2):276. doi: 10.3390/biom15020276. Biomolecules. 2025. PMID: 40001580 Free PMC article.
-
NSD2-mediated H3K36me2 exacerbates osteoporosis via activation of hoxa2 in bone marrow mesenchymal stem cells.Cell Signal. 2024 Sep;121:111294. doi: 10.1016/j.cellsig.2024.111294. Epub 2024 Jul 10. Cell Signal. 2024. PMID: 38996954
-
Melatonin activates mitochondrial unfolded protein response to preserve osteogenic potential of senescent BMSCs via upregulating PDI-6.Biochimie. 2023 Jun;209:44-51. doi: 10.1016/j.biochi.2023.01.015. Epub 2023 Jan 26. Biochimie. 2023. PMID: 36708867
-
NAP1L2 drives mesenchymal stem cell senescence and suppresses osteogenic differentiation.Aging Cell. 2022 Feb;21(2):e13551. doi: 10.1111/acel.13551. Epub 2022 Jan 15. Aging Cell. 2022. PMID: 35032339 Free PMC article.
-
Drug Discovery Targeting Nuclear Receptor Binding SET Domain Protein 2 (NSD2).J Med Chem. 2023 Aug 24;66(16):10991-11026. doi: 10.1021/acs.jmedchem.3c00948. Epub 2023 Aug 14. J Med Chem. 2023. PMID: 37578463 Free PMC article. Review.
Cited by
-
Age-related alveolar bone maladaptation in adult orthodontics: finding new ways out.Int J Oral Sci. 2024 Aug 1;16(1):52. doi: 10.1038/s41368-024-00319-7. Int J Oral Sci. 2024. PMID: 39085217 Free PMC article. Review.
-
Mechanisms during Osteogenic Differentiation in Human Dental Follicle Cells.Int J Mol Sci. 2022 May 25;23(11):5945. doi: 10.3390/ijms23115945. Int J Mol Sci. 2022. PMID: 35682637 Free PMC article. Review.
-
Rejuvenation of Bone Marrow Mesenchymal Stem Cells: Mechanisms and Their Application in Senile Osteoporosis Treatment.Biomolecules. 2025 Feb 13;15(2):276. doi: 10.3390/biom15020276. Biomolecules. 2025. PMID: 40001580 Free PMC article.
-
Dysregulation of histone modifications in bone marrow mesenchymal stem cells during skeletal ageing: roles and therapeutic prospects.Stem Cell Res Ther. 2023 Jun 25;14(1):166. doi: 10.1186/s13287-023-03393-6. Stem Cell Res Ther. 2023. PMID: 37357311 Free PMC article. Review.
-
Antisenescence therapies for age-related bone loss: Target factors, medicines, biomedical materials.Clin Transl Med. 2025 Jun;15(6):e70350. doi: 10.1002/ctm2.70350. Clin Transl Med. 2025. PMID: 40490937 Free PMC article. Review.
References
-
- Pevet P. Melatonin and biological rhythms. Therapie. 1998;53:411‐420. - PubMed
-
- Radio NM, Doctor JS, Witt‐Enderby PA. Melatonin enhances alkaline phosphatase activity in differentiating human adult mesenchymal stem cells grown in osteogenic medium via MT2 melatonin receptors and the MEK/ERK (1/2) signaling cascade. J Pineal Res. 2006;40:332‐342. 10.1111/j.1600-079X.2006.00318.x - DOI - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- 81870161/National Natural Science Foundation of China
- 82070221/National Natural Science Foundation of China
- 81900215/National Natural Science Foundation of China
- 81471175/National Natural Science Foundation of China
- Z200020/Beijing Natural Science Foundation of China
- 2019QL009/Academic promotion programme of Shandong First Medical University
- 201810062011/College Student Innovation Program of Tianjin Medical University
- 202010062030/College Student Innovation Program of Tianjin Medical University
- 202110062001/College Student Innovation Program of Tianjin Medical University
- 2019YJSB110/Tianjin Research Innovation Project for Postgraduate Students
- 2020YJSB162/Tianjin Research Innovation Project for Postgraduate Students
LinkOut - more resources
Full Text Sources