

LAP2α preserves genome integrity through assisting RPA deposition on damaged chromatin
- PMID: 35227284
- PMCID: PMC8883701
- DOI: 10.1186/s13059-022-02638-6
LAP2α preserves genome integrity through assisting RPA deposition on damaged chromatin
Abstract
Background: Single-stranded DNA (ssDNA) coated with replication protein A (RPA) acts as a key platform for the recruitment and exchange of genome maintenance factors in DNA damage response. Yet, how the formation of the ssDNA-RPA intermediate is regulated remains elusive.
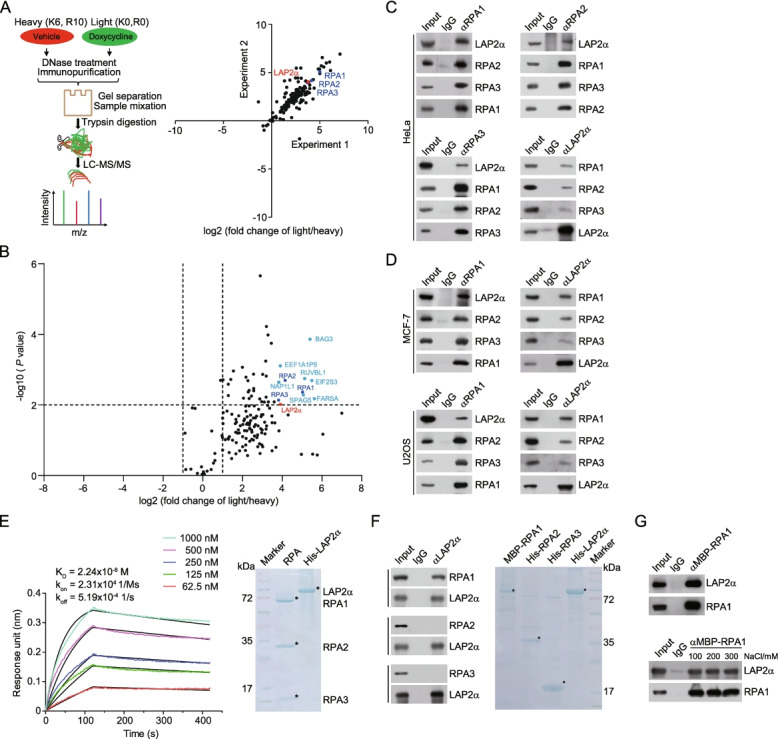
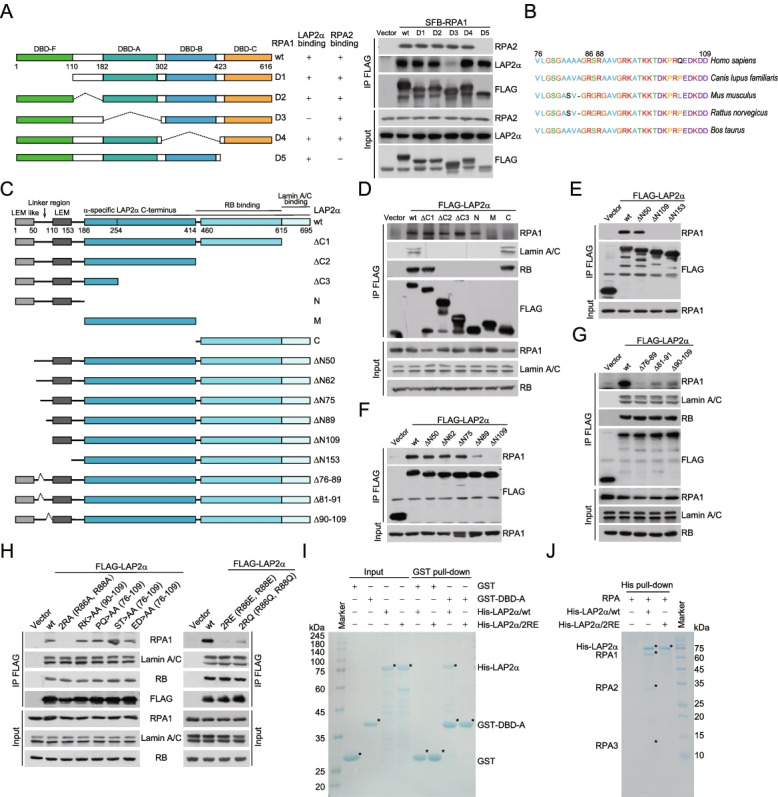
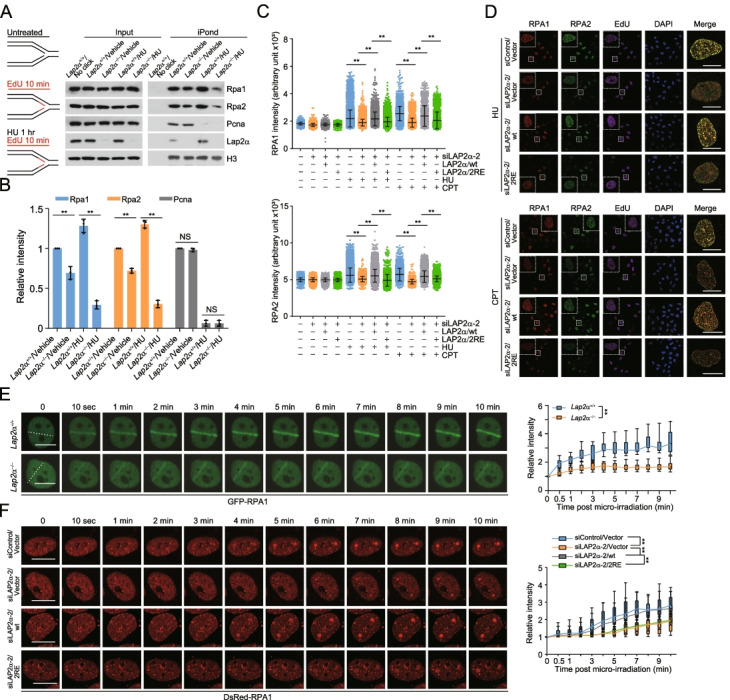
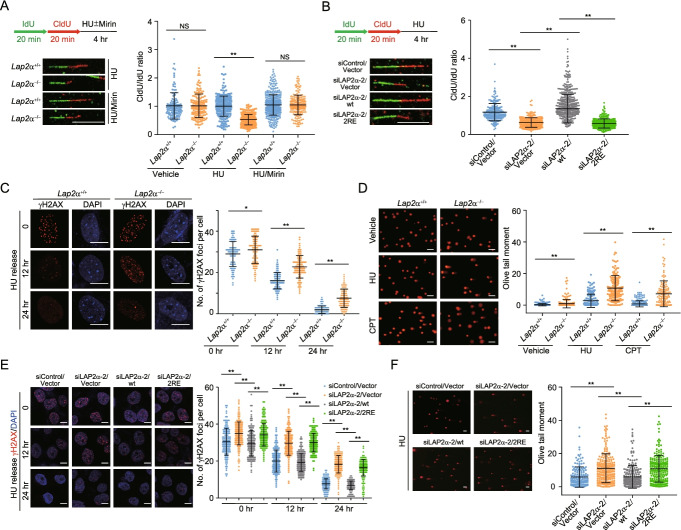
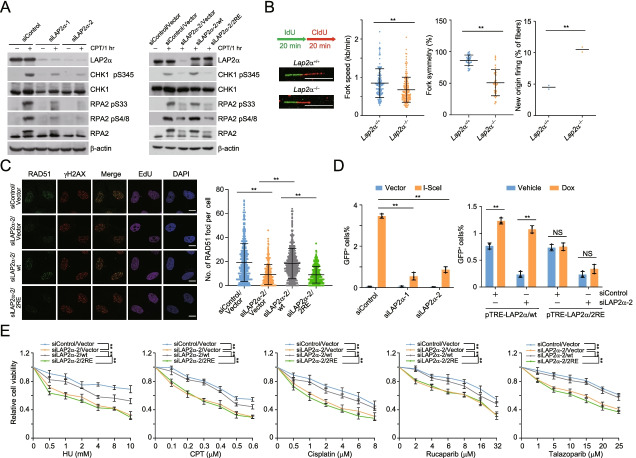
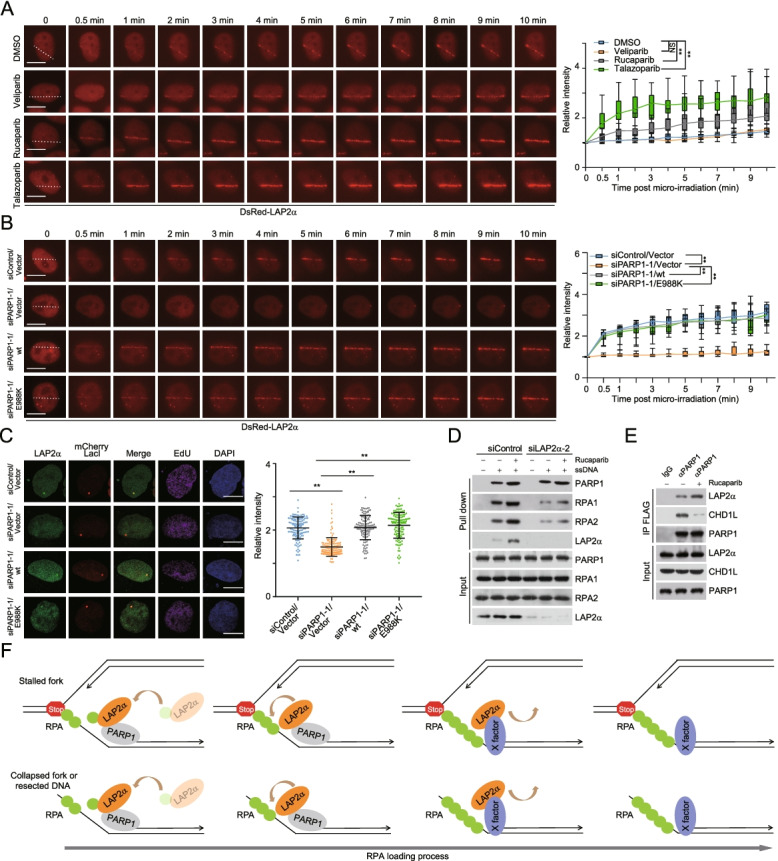
Results: Here, we report that the lamin-associated protein LAP2α is physically associated with RPA, and LAP2α preferentially facilitates RPA deposition on damaged chromatin via physical contacts between LAP2α and RPA1. Importantly, LAP2α-promoted RPA binding to ssDNA plays a critical role in protection of replication forks, activation of ATR, and repair of damaged DNA. We further demonstrate that the preference of LAP2α-promoted RPA loading on damaged chromatin depends on poly ADP-ribose polymerase PARP1, but not poly(ADP-ribosyl)ation.
Conclusions: Our study provides mechanistic insight into RPA deposition in response to DNA damage and reveals a genome protection role of LAP2α.
Keywords: ATR activation; Genome stability; Homologous recombination; LAP2α; PARP1; RPA loading.
© 2022. The Author(s).
Conflict of interest statement
The authors declare that they have no competing interests.
Figures
References
-
- Blackford AN, Jackson SP. ATM, ATR, and DNA-PK: the trinity at the heart of the DNA damage response. Mol Cell. 2017;66:801–817. - PubMed
-
- Halazonetis TD, Gorgoulis VG, Bartek J. An oncogene-induced DNA damage model for cancer development. Science. 2008;319:1352–1355. - PubMed
-
- Bartkova J, Horejsi Z, Koed K, Kramer A, Tort F, Zieger K, et al. DNA damage response as a candidate anti-cancer barrier in early human tumorigenesis. Nature. 2005;434:864–870. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
