

Polarized epithelium-sperm co-culture system reveals stimulatory factors for the secretion of mouse epididymal quiescin sulfhydryl oxidase 1
- PMID: 35228412
- PMCID: PMC9184822
- DOI: 10.1262/jrd.2021-128
Polarized epithelium-sperm co-culture system reveals stimulatory factors for the secretion of mouse epididymal quiescin sulfhydryl oxidase 1
Abstract
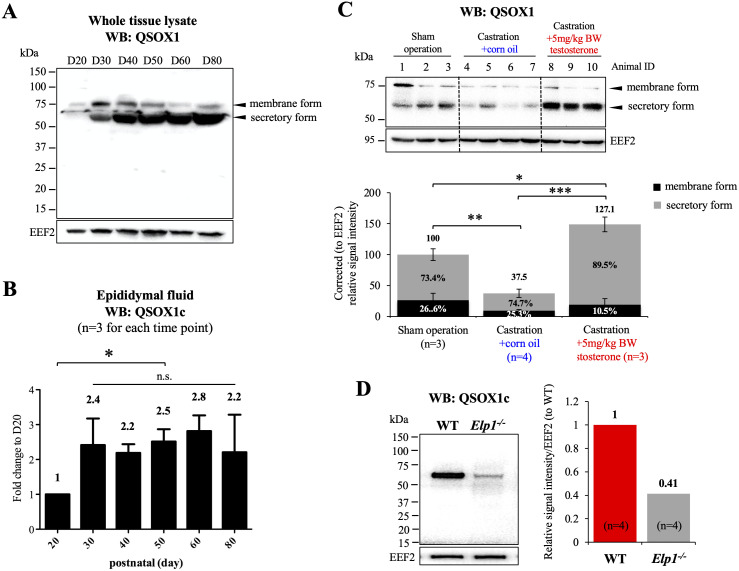
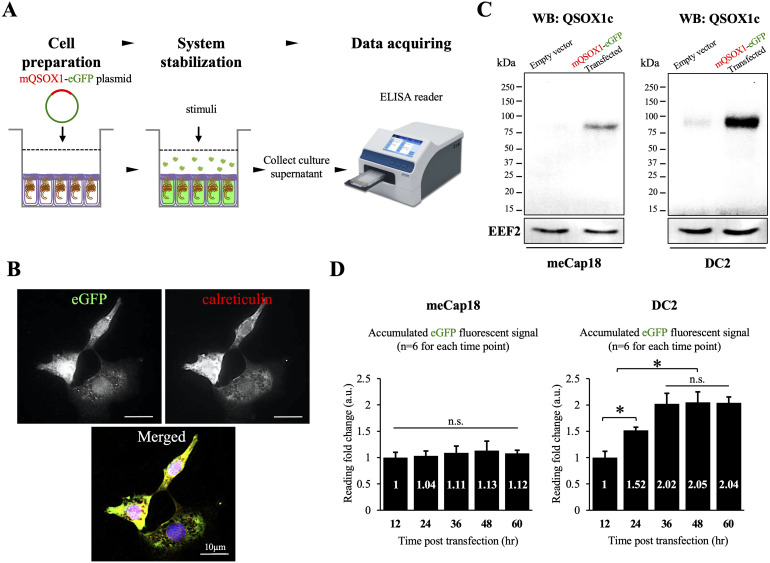
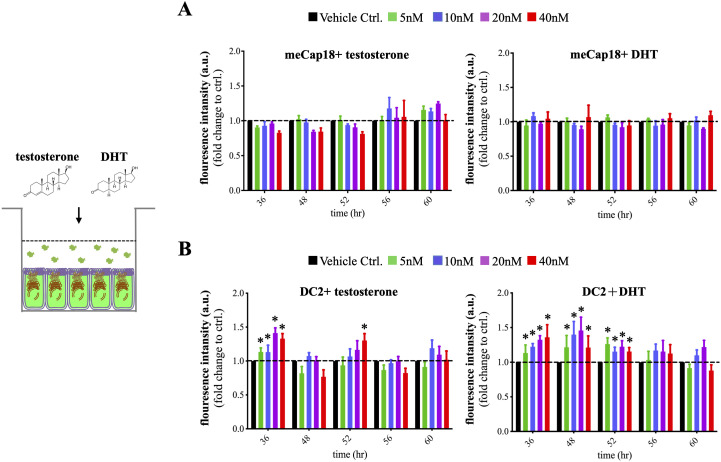
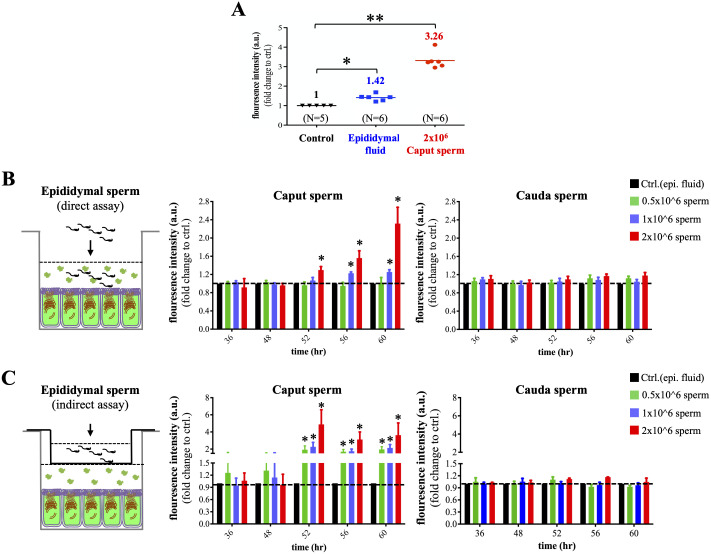
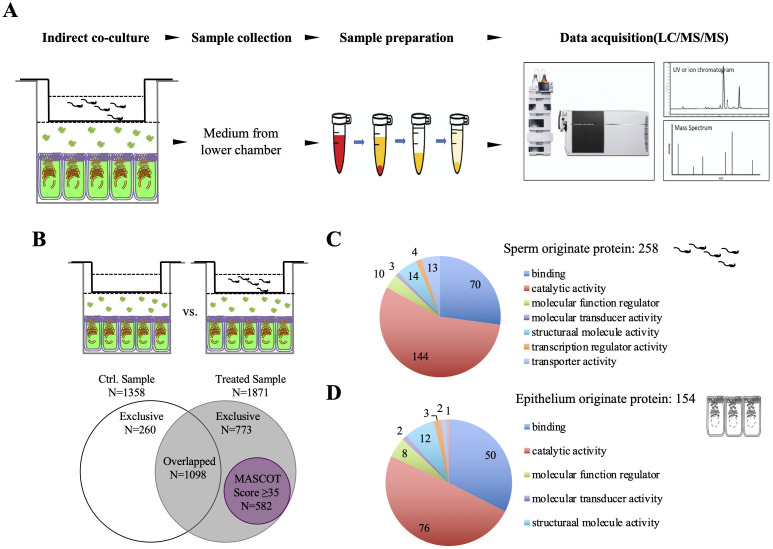
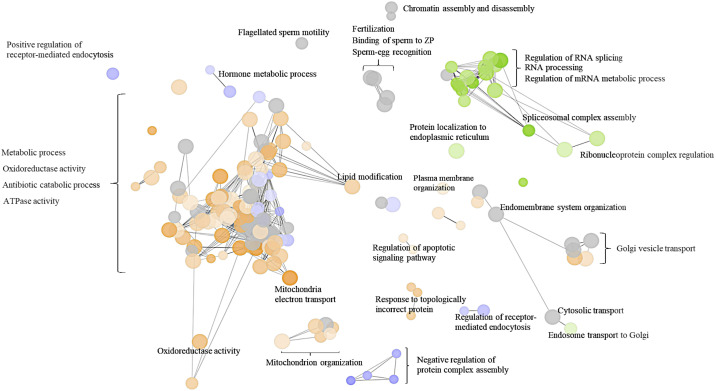
Spermatozoa acquire fertilization ability through post-translational modifications. These membrane surface alterations occur in various segments of the epididymis. Quiescin sulfhydryl oxidases, which catalyze thiol-oxidation reactions, are involved in disulfide bond formation, which is essential for sperm maturation, upon transition and migration in the epididymis. Using castration and azoospermia transgenic mouse models, in the present study, we showed that quiescin sulfhydryl oxidase 1 (QSOX1) protein expression and secretion are positively correlated with the presence of testosterone and sperm cells. A two-dimensional in vitro epithelium-sperm co-culture system provided further evidence in support of the notion that both testosterone and its dominant metabolite, 5α-dihydrotestosterone, promote epididymal QSOX1 secretion. We also demonstrated that immature caput spermatozoa, but not mature cauda sperm cells, exhibited great potential to stimulate QSOX1 secretion in vitro, suggesting that sperm maturation is a key regulatory factor for mouse epididymal QSOX1 secretion. Proteomic analysis identified 582 secretory proteins from the co-culture supernatant, of which 258 were sperm-specific and 154 were of epididymal epithelium-origin. Gene Ontology analysis indicated that these secreted proteins exhibit functions known to facilitate sperm membrane organization, cellular activity, and sperm-egg recognition. Taken together, our data demonstrated that testosterone and sperm maturation status are key regulators of mouse epididymal QSOX1 protein expression and secretion.
Keywords: Epididymis; Epithelium; Fertility; Quiescin sulfhydryl oxidase; Spermatozoa.
Conflict of interest statement
The authors declare no conflicts of interest that could prejudice the impartiality of the research reported.
Figures
Similar articles
-
Mouse quiescin sulfhydryl oxidases exhibit distinct epididymal luminal distribution with segment-specific sperm surface associations.Biol Reprod. 2018 Nov 1;99(5):1022-1033. doi: 10.1093/biolre/ioy125. Biol Reprod. 2018. PMID: 29800099
-
Testosterone regulation on quiescin sulfhydryl oxidase 2 synthesis in the epididymis.Reproduction. 2021 Apr 20;161(5):593-602. doi: 10.1530/REP-20-0629. Reproduction. 2021. PMID: 33784244
-
Junctional adhesion molecule A: expression in the murine epididymal tract and accessory organs and acquisition by maturing sperm.Mol Hum Reprod. 2017 Feb 10;23(2):132-140. doi: 10.1093/molehr/gaw082. Mol Hum Reprod. 2017. PMID: 28062807 Free PMC article.
-
Role of exosomes in sperm maturation during the transit along the male reproductive tract.Blood Cells Mol Dis. 2005 Jul-Aug;35(1):1-10. doi: 10.1016/j.bcmd.2005.03.005. Blood Cells Mol Dis. 2005. PMID: 15893944 Review.
-
The current perspective on genetic and epigenetic factors in sperm maturation in the epididymis.Andrologia. 2021 Apr;53(3):e13989. doi: 10.1111/and.13989. Epub 2021 Jan 25. Andrologia. 2021. PMID: 33491190 Review.
Cited by
-
Advancement and Potential Applications of Epididymal Organoids.Biomolecules. 2024 Aug 17;14(8):1026. doi: 10.3390/biom14081026. Biomolecules. 2024. PMID: 39199413 Free PMC article. Review.
-
Improvement of Fertilization Capacity and Developmental Ability of Vitrified Bovine Oocytes by JUNO mRNA Microinjection and Cholesterol-Loaded Methyl-β-Cyclodextrin Treatment.Int J Mol Sci. 2022 Dec 29;24(1):590. doi: 10.3390/ijms24010590. Int J Mol Sci. 2022. PMID: 36614032 Free PMC article.
References
-
- Nixon B, De Iuliis GN, Hart HM, Zhou W, Mathe A, Bernstein IR, Anderson AL, Stanger SJ, Skerrett-Byrne DA, Jamaluddin MFB, Almazi JG, Bromfield EG, Larsen MR, Dun MD. Proteomic Profiling of Mouse Epididymosomes Reveals their Contributions to Post-testicular Sperm Maturation. Mol Cell Proteomics 2019; 18(Suppl 1): S91–S108. - PMC - PubMed
-
- Sipilä P, Björkgren I. Segment-specific regulation of epididymal gene expression. Reproduction 2016; 152: R91–R99. - PubMed
-
- Sullivan R, Légaré C, Lamontagne-Proulx J, Breton S, Soulet D. Revisiting structure/functions of the human epididymis. Andrology 2019; 7: 748–757. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous