

Squalene epoxidase drives cancer cell proliferation and promotes gut dysbiosis to accelerate colorectal carcinogenesis
- PMID: 35232776
- PMCID: PMC9554078
- DOI: 10.1136/gutjnl-2021-325851
Squalene epoxidase drives cancer cell proliferation and promotes gut dysbiosis to accelerate colorectal carcinogenesis
Abstract
Objective: Aberrant lipid metabolism is a hallmark of colorectal cancer (CRC). Squalene epoxidase (SQLE), a rate-limiting enzyme in cholesterol biosynthesis, is upregulated in CRC. Here, we aim to determine oncogenic function of SQLE and its interplay with gut microbiota in promoting colorectal tumourigenesis.
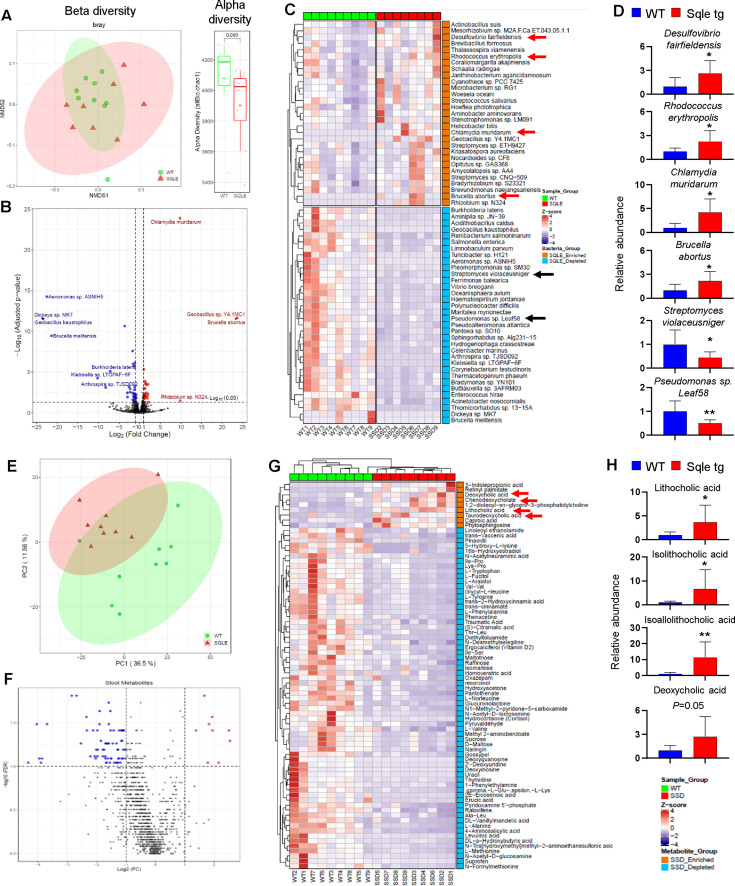
Design: Paired adjacent normal tissues and CRC from two cohorts were analysed (n=202). Colon-specific Sqle transgenic (Sqle tg) mice were generated by crossing Rosa26-lsl-Sqle mice to Cdx2-Cre mice. Stools were collected for metagenomic and metabolomic analyses.
Results: SQLE messenger RNA and protein expression was upregulated in CRC (p<0.01) and predict poor survival of patients with CRC. SQLE promoted CRC cell proliferation by inducing cell cycle progression and suppressing apoptosis. In azoxymethane-induced CRC model, Sqle tg mice showed increased tumourigenesis compared with wild-type mice (p<0.01). Integrative metagenomic and metabolomic analyses unveiled gut dysbiosis in Sqle tg mice with enriched pathogenic bacteria, which was correlated to increased secondary bile acids. Consistent with detrimental effect of secondary bile acids, gut barrier function was impaired in Sqle tg mice, with reduced tight junction proteins Jam-c and occludin. Transplantation of Sqle tg mice stool to germ-free mice impaired gut barrier function and stimulated cell proliferation compared with control mice stool. Finally, we demonstrated that terbinafine, a SQLE inhibitor, could be repurposed for CRC by synergising with oxaliplatin and 5-fluorouracil to inhibit CRC growth.
Conclusion: This study demonstrates that SQLE mediates oncogenesis via cell intrinsic effects and modulation of gut microbiota-metabolite axis. SQLE represents a therapeutic target and prognostic marker in CRC.
Keywords: colonic microflora; colorectal cancer; colorectal neoplasm.
© Author(s) (or their employer(s)) 2022. Re-use permitted under CC BY-NC. No commercial re-use. See rights and permissions. Published by BMJ.
Conflict of interest statement
Competing interests: None declared.
Figures






References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Miscellaneous