

Structural basis for mismatch surveillance by CRISPR-Cas9
- PMID: 35236982
- PMCID: PMC8907077
- DOI: 10.1038/s41586-022-04470-1
Structural basis for mismatch surveillance by CRISPR-Cas9
Erratum in
-
Publisher Correction: Structural basis for mismatch surveillance by CRISPR-Cas9.Nature. 2022 Apr;604(7904):E10. doi: 10.1038/s41586-022-04655-8. Nature. 2022. PMID: 35319014 Free PMC article. No abstract available.
Abstract
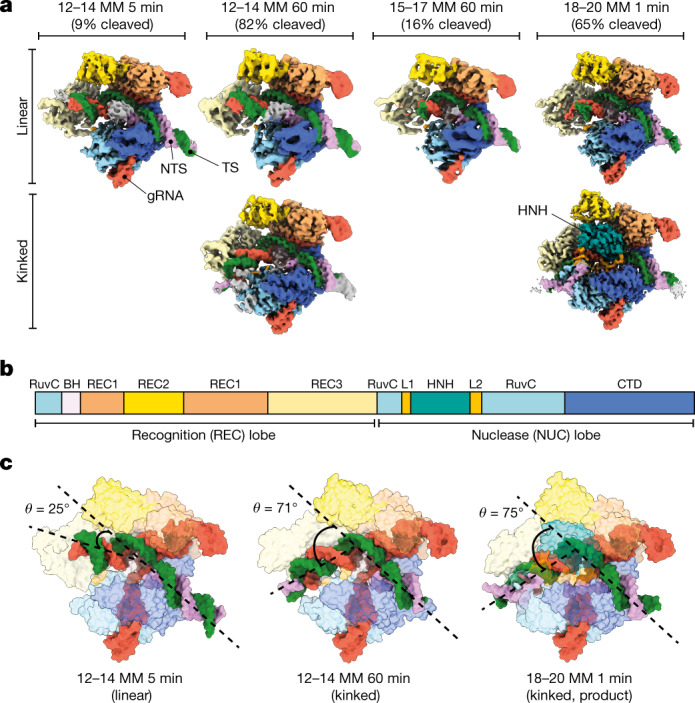
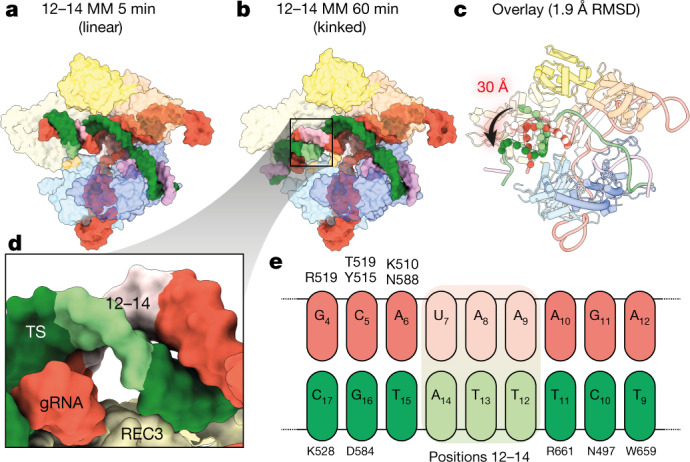
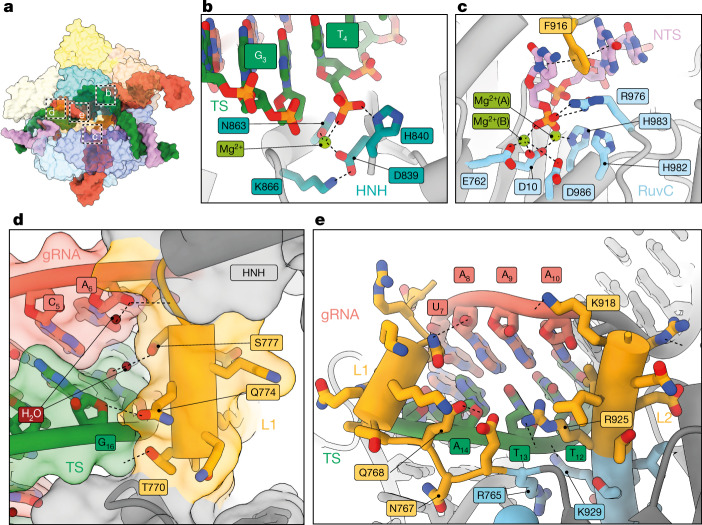
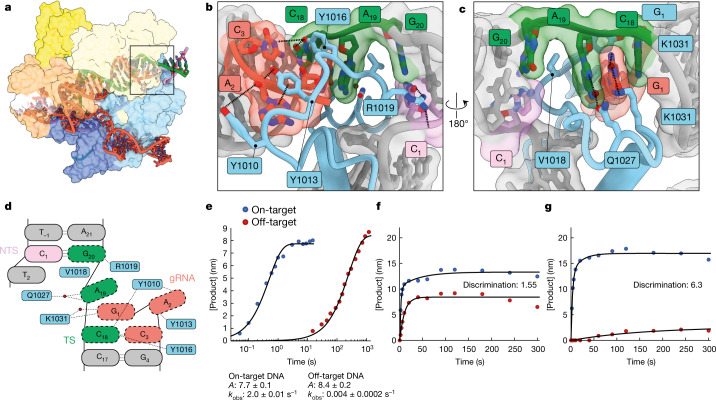
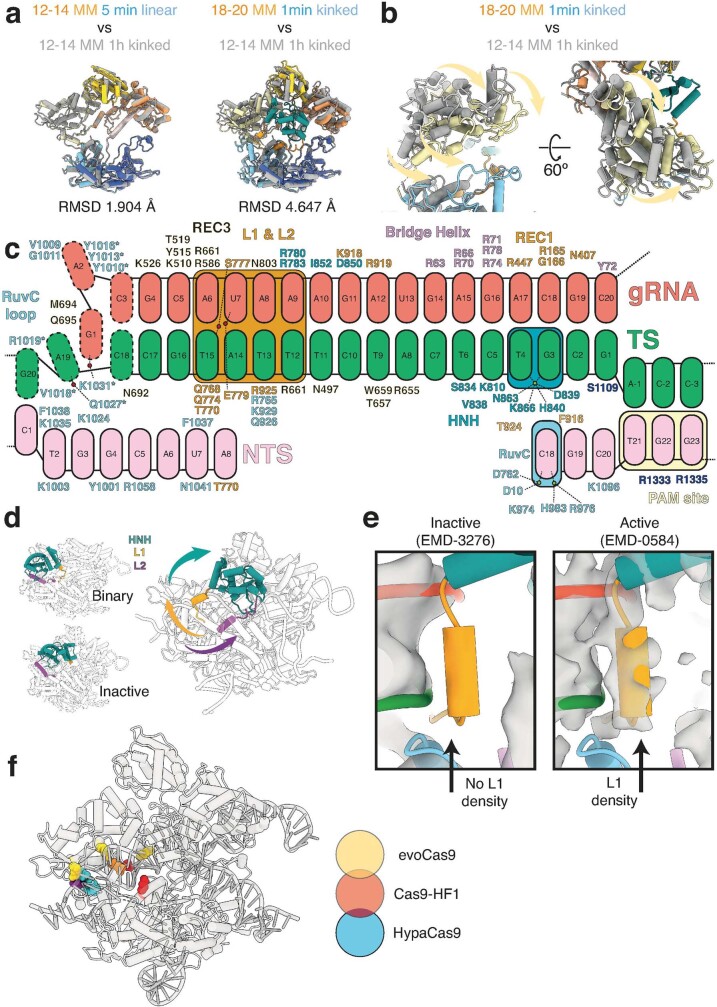
CRISPR-Cas9 as a programmable genome editing tool is hindered by off-target DNA cleavage1-4, and the underlying mechanisms by which Cas9 recognizes mismatches are poorly understood5-7. Although Cas9 variants with greater discrimination against mismatches have been designed8-10, these suffer from substantially reduced rates of on-target DNA cleavage5,11. Here we used kinetics-guided cryo-electron microscopy to determine the structure of Cas9 at different stages of mismatch cleavage. We observed a distinct, linear conformation of the guide RNA-DNA duplex formed in the presence of mismatches, which prevents Cas9 activation. Although the canonical kinked guide RNA-DNA duplex conformation facilitates DNA cleavage, we observe that substrates that contain mismatches distal to the protospacer adjacent motif are stabilized by reorganization of a loop in the RuvC domain. Mutagenesis of mismatch-stabilizing residues reduces off-target DNA cleavage but maintains rapid on-target DNA cleavage. By targeting regions that are exclusively involved in mismatch tolerance, we provide a proof of concept for the design of next-generation high-fidelity Cas9 variants.
© 2022. The Author(s).
Conflict of interest statement
J.P.K.B., M.-S.L., G.N.H., T.L.D., K.A.J. and D.W.T. are inventors on a patent application based on this research titled ‘Methods and compositions for improved Cas9 specificity’ filed by the Board of Regents, The University of Texas System. The US Patent and Trademark Office (USPTO) has assigned US application no. 63/243,481 to this application, and the filing date of 13 September 2021. K.A.J. is the president of KinTek, which provided the chemical-quench flow instruments and the KinTek Explorer software used in this study.
Figures
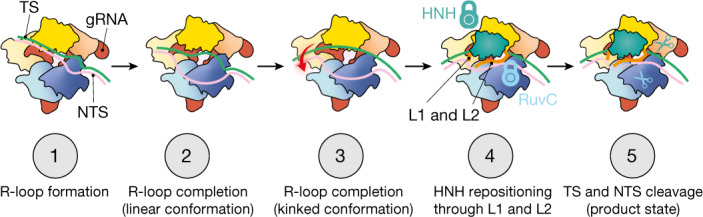
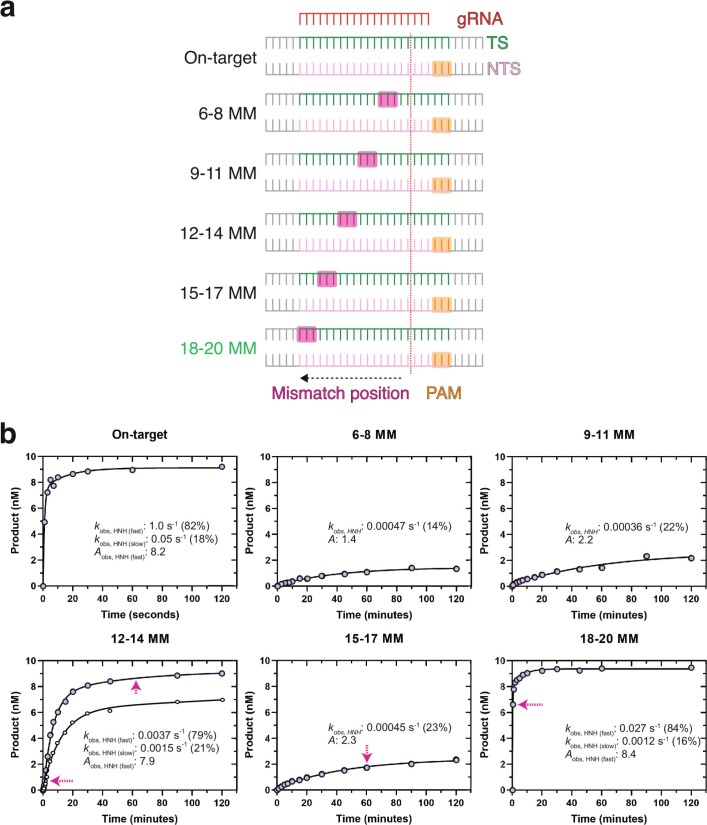
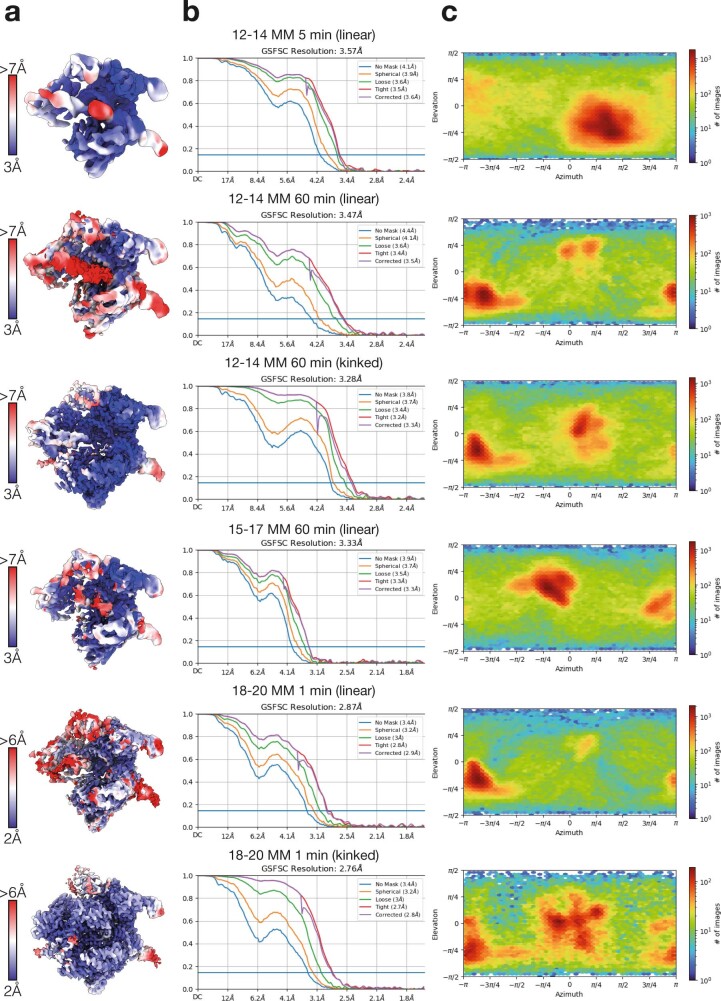
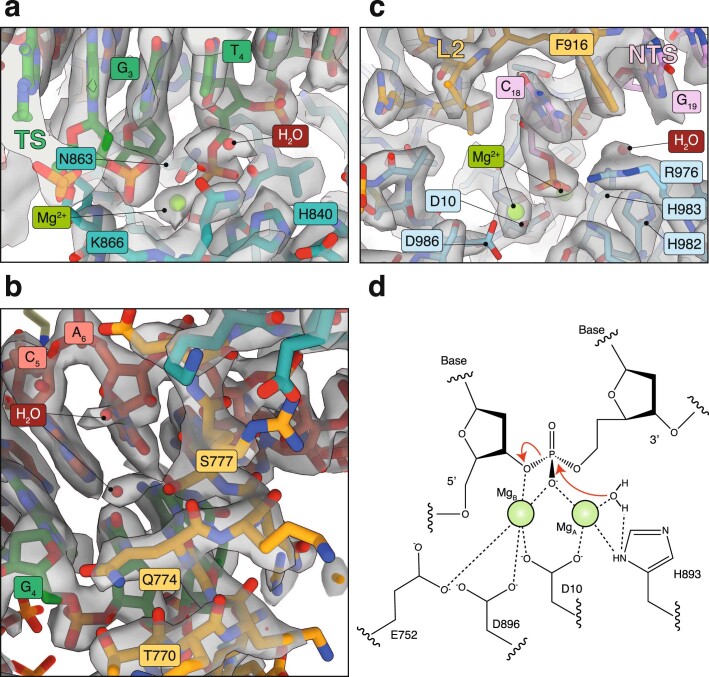
Comment in
-
SuperFi-Cas9: High Fidelity Meets High Activity.CRISPR J. 2022 Apr;5(2):171-173. doi: 10.1089/crispr.2022.29146.ywa. CRISPR J. 2022. PMID: 35438514 No abstract available.
-
Structural insights into Cas9 mismatch: promising for development of high-fidelity Cas9 variants.Signal Transduct Target Ther. 2022 Aug 4;7(1):271. doi: 10.1038/s41392-022-01139-z. Signal Transduct Target Ther. 2022. PMID: 35927241 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
