

Orientin Prolongs the Longevity of Caenorhabditis elegans and Postpones the Development of Neurodegenerative Diseases via Nutrition Sensing and Cellular Protective Pathways
- PMID: 35237385
- PMCID: PMC8885179
- DOI: 10.1155/2022/8878923
Orientin Prolongs the Longevity of Caenorhabditis elegans and Postpones the Development of Neurodegenerative Diseases via Nutrition Sensing and Cellular Protective Pathways
Abstract
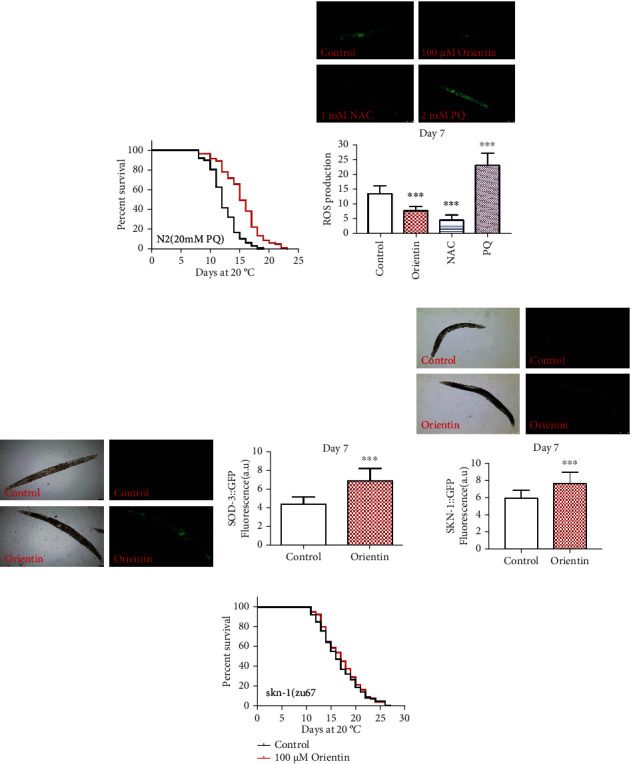
Age is the major risk factor for most of the deadliest diseases. Developing small molecule drugs with antiaging effects could improve the health of aged people and retard the onset and progress of aging-associated disorders. Bioactive secondary metabolites from medicinal plants are the main source for development of medication. Orientin is a water-soluble flavonoid monomer compound widely found in many medicinal plants. Orientin inhibits fat production, antioxidation, and anti-inflammatory activities. In this study, we explored whether orientin could affect the aging of C. elegans. We found that orientin improved heat, oxidative, and pathogenic stress resistances through activating stress responses, including HSF-1-mediated heat shock response, SKN-1-mediated xenobiotic and oxidation response, mitochondria unfolded responses, endoplasmic unfolded protein response, and increased autophagy activity. Orientin also could activate key regulators of the nutrient sensing pathway, including AMPK and insulin downstream transcription factor FOXO/DAF-16 to further improve the cellular health status. The above effects of orientin reduced the accumulation of toxic proteins (α-synuclein, β-amyloid, and poly-Q) and delayed the onset of neurodegenerative disorders in AD, PD, and HD models of C. elegans and finally increased the longevity and health span of C. elegans. Our results suggest that orientin has promising antiaging effects and could be a potential natural source for developing novel therapeutic drugs for aging and its related diseases.
Copyright © 2022 Yuan Qu et al.
Conflict of interest statement
The authors declare that there is no conflict of interest.
Figures








References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Research Materials
Miscellaneous
