

Efficient polymer nanoparticle-mediated delivery of gene editing reagents into human hematopoietic stem and progenitor cells
- PMID: 35240320
- PMCID: PMC9171380
- DOI: 10.1016/j.ymthe.2022.02.026
Efficient polymer nanoparticle-mediated delivery of gene editing reagents into human hematopoietic stem and progenitor cells
Abstract
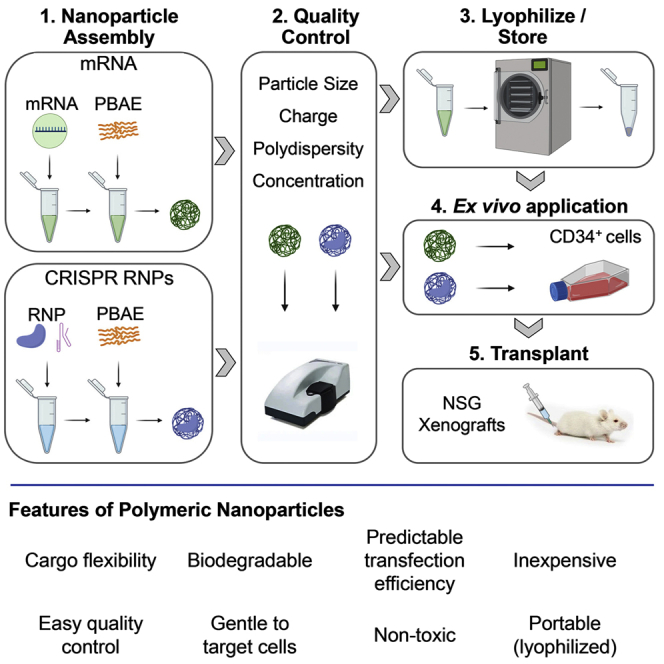
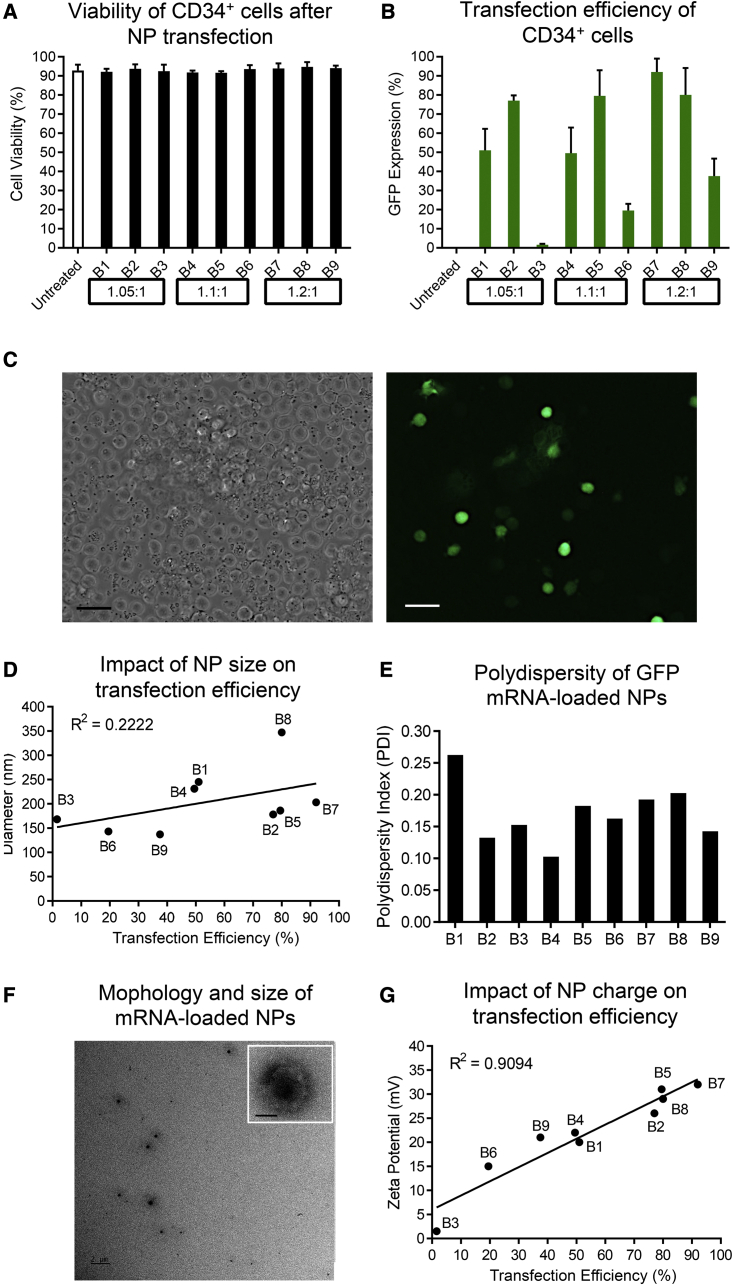
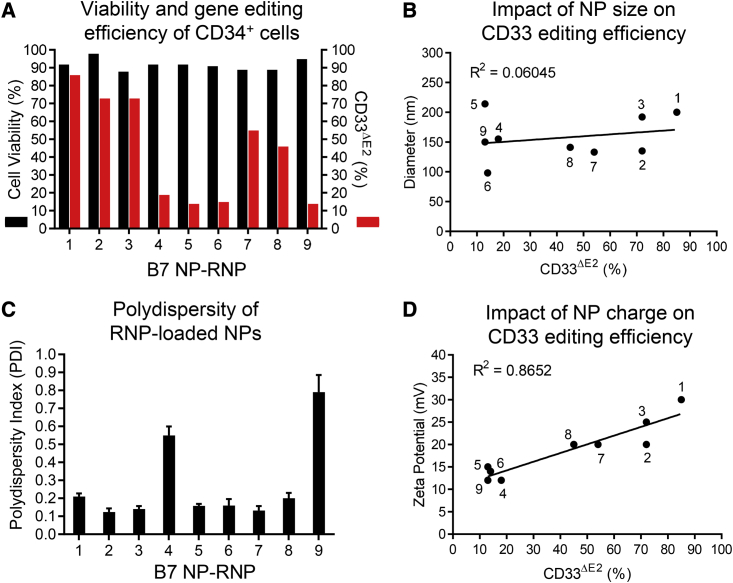
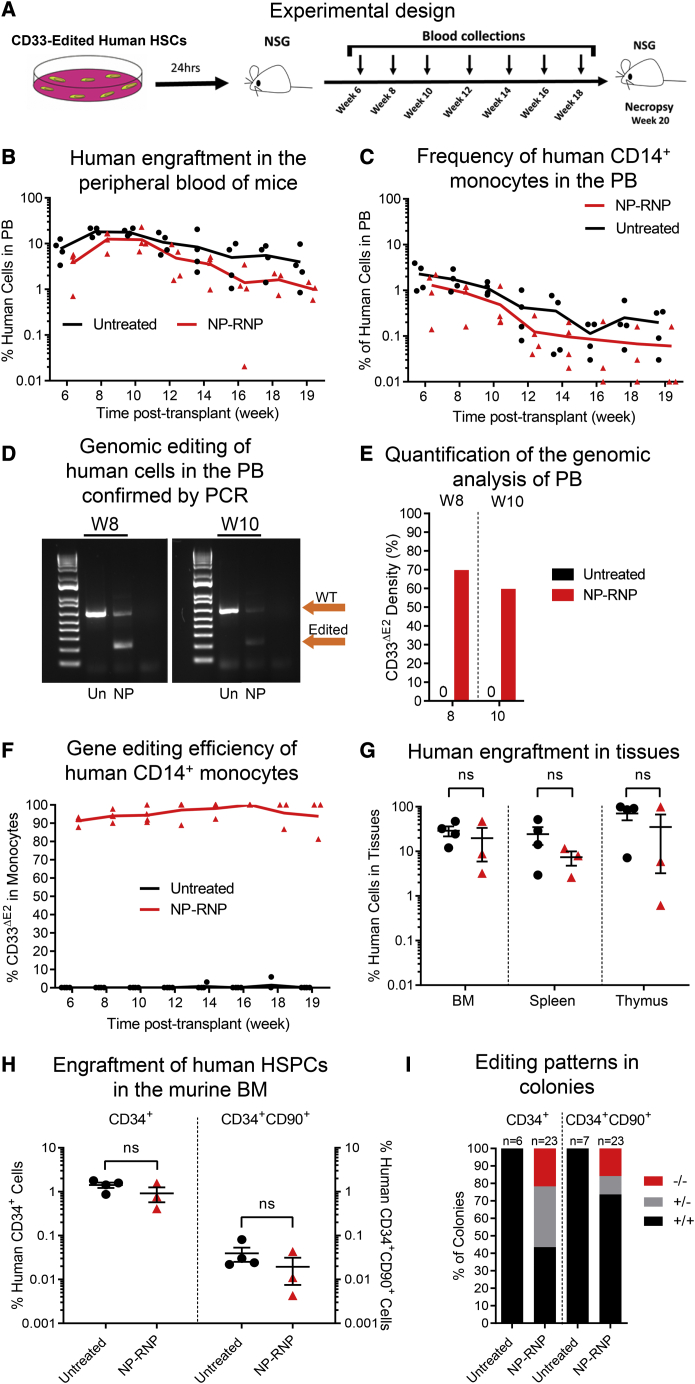
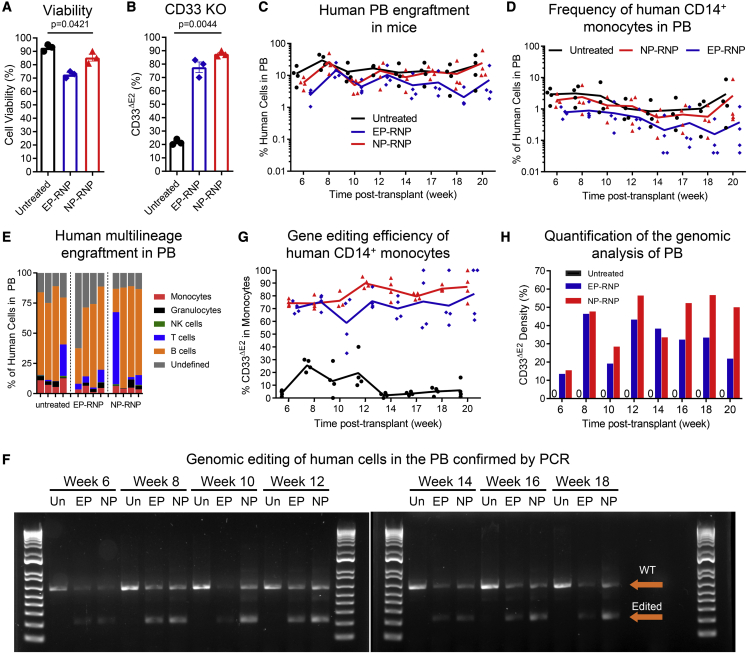
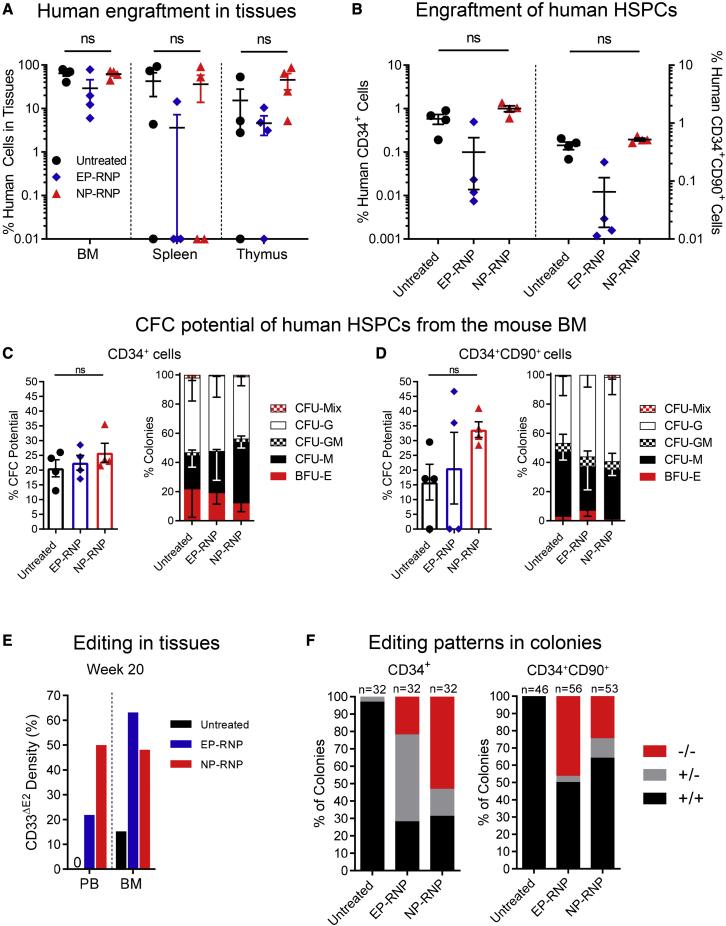
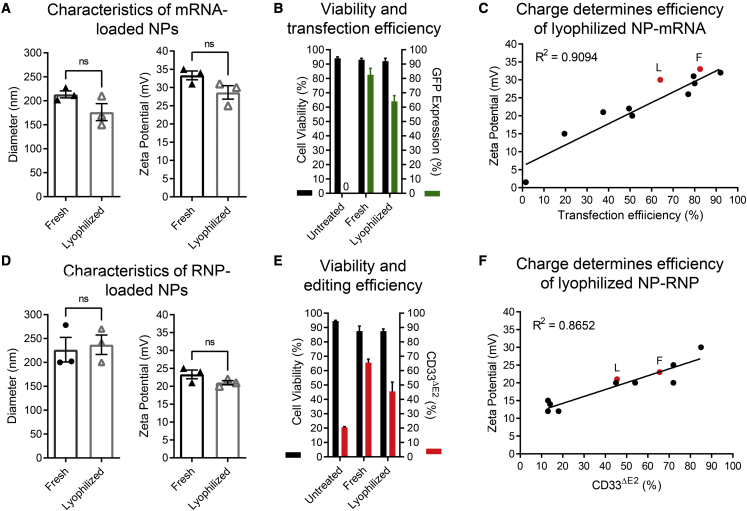
Clinical applications of hematopoietic stem cell (HSC) gene editing are limited due to their complex and expensive logistics. HSC editing is commonly performed ex vivo using electroporation and requires good manufacturing practice (GMP) facilities, similar to bone marrow transplant centers. In vivo gene editing could overcome this limitation; however, electroporation is unsuitable for systemic in vivo applications to HSCs. Here we evaluated polymer-based nanoparticles (NPs), which could also be used for in vivo administration, for the delivery of mRNA and nucleases to human granulocyte colony-stimulating factor (GCSF)-mobilized CD34+ cells. NP-mediated ex vivo delivery showed no toxicity, and the efficiency was directly correlated with the charge of the NPs. In a side-by-side comparison with electroporation, NP-mediated gene editing allowed for a 3-fold reduction in the amount of reagents, with similar efficiency. Furthermore, we observed enhanced engraftment potential of human HSCs in the NSG mouse xenograft model using NPs. Finally, mRNA- and nuclease-loaded NPs were successfully lyophilized for storage, maintaining their transfection potential after rehydration. In conclusion, we show that polymer-based NP delivery of mRNA and nucleases has the potential to overcome current limitations of HSC gene editing. The predictable transfection efficiency, low toxicity, and ability to lyophilize NPs will greatly enhance the portability and provide a highly promising platform for HSC gene therapy.
Keywords: gene editing nucleases; gene therapy; hematopoietic stem cells; lyophilization; polymeric nanoparticles; quality control standards.
Copyright © 2022. Published by Elsevier Inc.
Conflict of interest statement
Declaration of interests S.R. is a consultant to Forty-Seven, Inc. (Gilead Sciences), and Ensoma, Inc. H.P.K. is or was a consultant to and has or had ownership interests with Rocket Pharmaceuticals, Homology Medicines, VOR Biopharma, and Ensoma, Inc. H.P.K. has also been a consultant to CSL Behring and Magenta Therapeutics.
Figures
References
-
- Naldini L. Gene therapy returns to centre stage (Review) Nature. 2015;526:351–360. - PubMed
-
- Frangoul H., Altshuler D., Cappellini M.D., Chen Y.S., Domm J., Eustace B.K., Foell J., de la Fuente J., Grupp S., Handgretinger R., et al. CRISPR-Cas9 cene editing for sickle cell disease and beta-thalassemia. N. Engl. J. Med. 2021;384:252–260. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Miscellaneous
