

ACSL4 contributes to ferroptosis-mediated rhabdomyolysis in exertional heat stroke
- PMID: 35243801
- PMCID: PMC9178366
- DOI: 10.1002/jcsm.12953
ACSL4 contributes to ferroptosis-mediated rhabdomyolysis in exertional heat stroke
Abstract
Background: Rhabdomyolysis (RM) is a common complication of exertional heat stroke (EHS) and constitutes a direct cause of death. However, the mechanism underlying RM following EHS remains unclear.
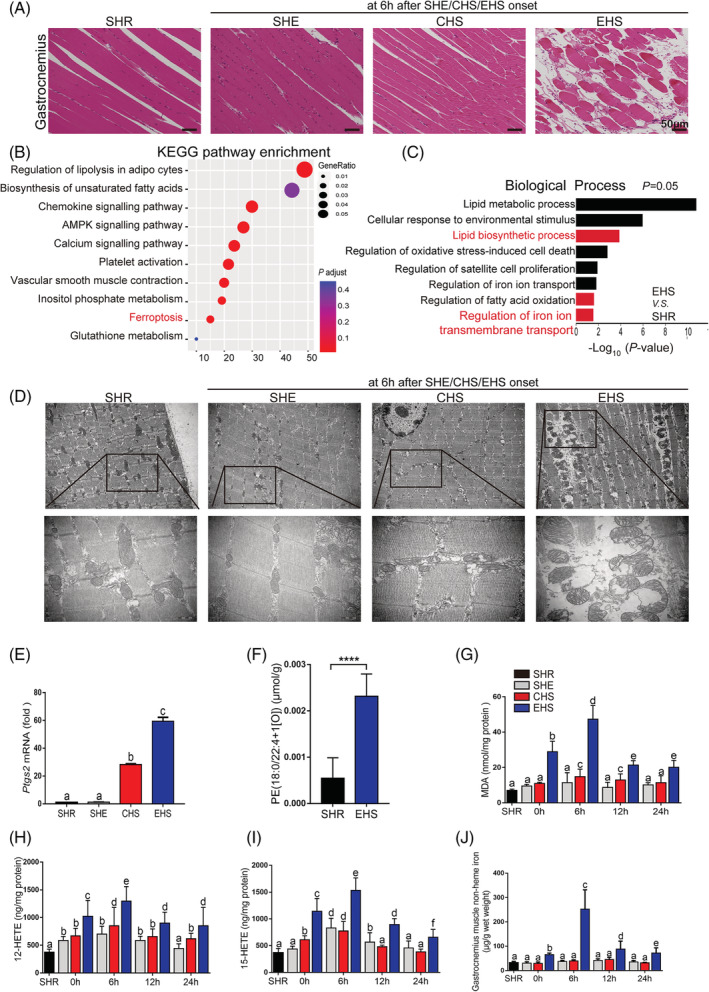
Methods: The murine EHS model was prepared by our previous protocol. RNA sequencing is applied to identify the pathological pathways that contribute to RM following EHS. Inhibition of the acyl-CoA synthetase long-chain family member 4 (ACSL4) was achieved by RNA silencing in vitro prior to ionomycin plus heat stress exposure or pharmacological inhibitors in vivo prior to heat and exertion exposure. The histological changes, the iron accumulation, oxidized phosphatidylethanolamines species, as well as histological evaluation and levels of lipid metabolites in skeletal muscle tissues were measured.
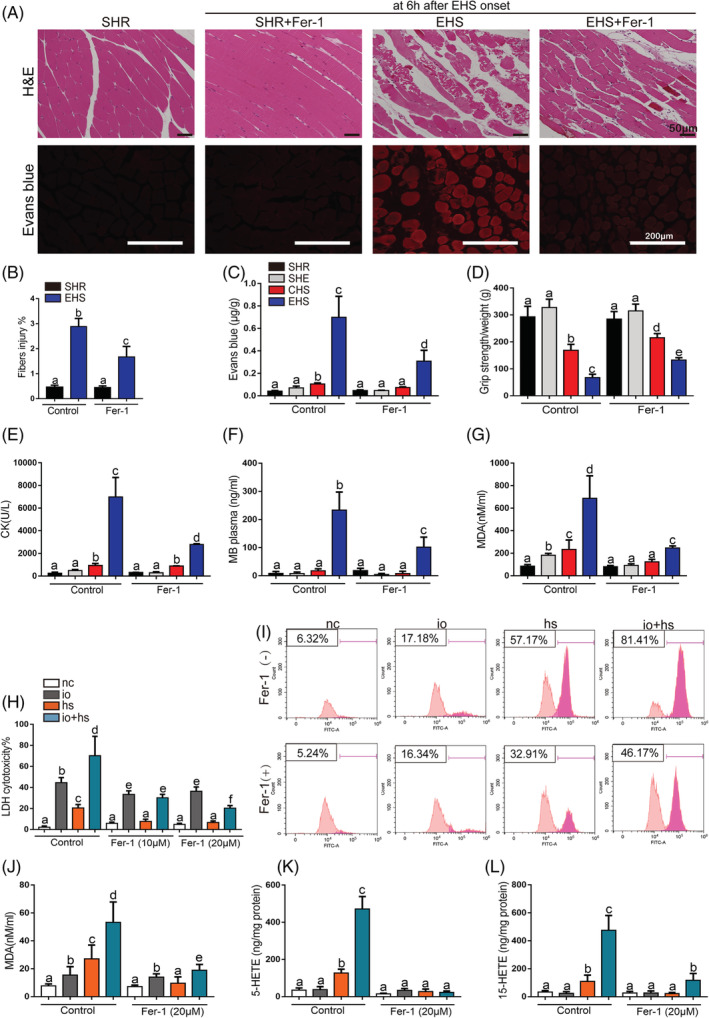
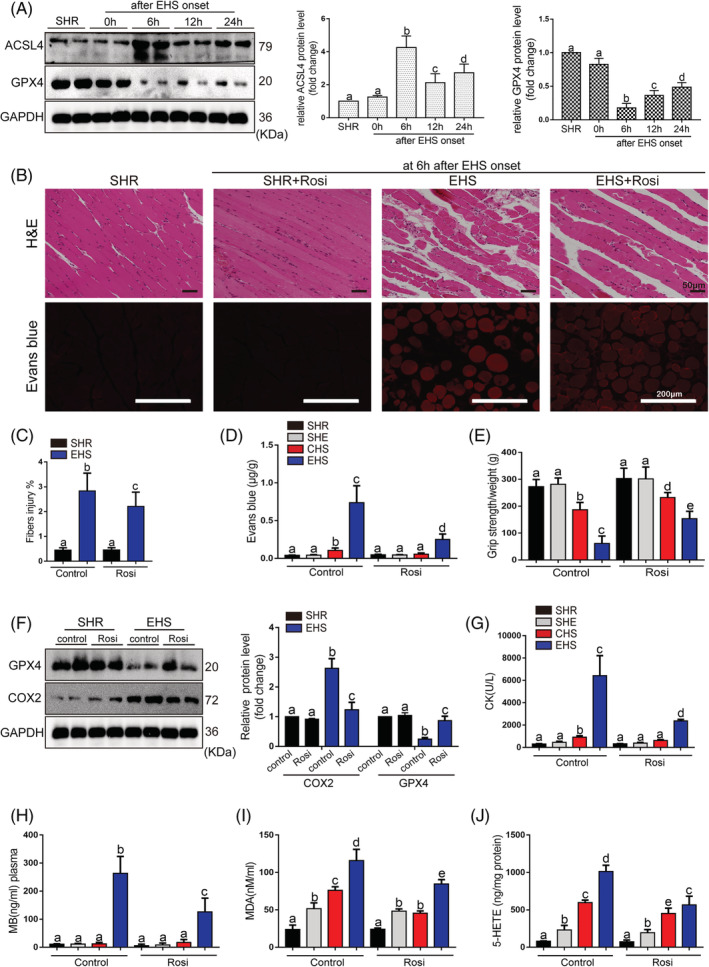
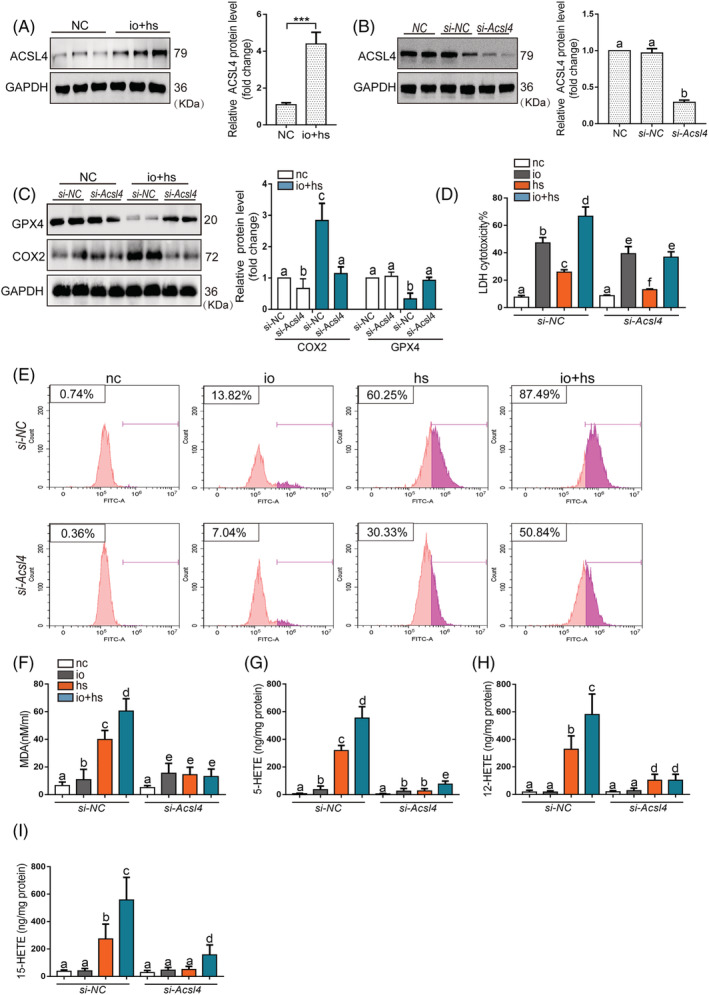
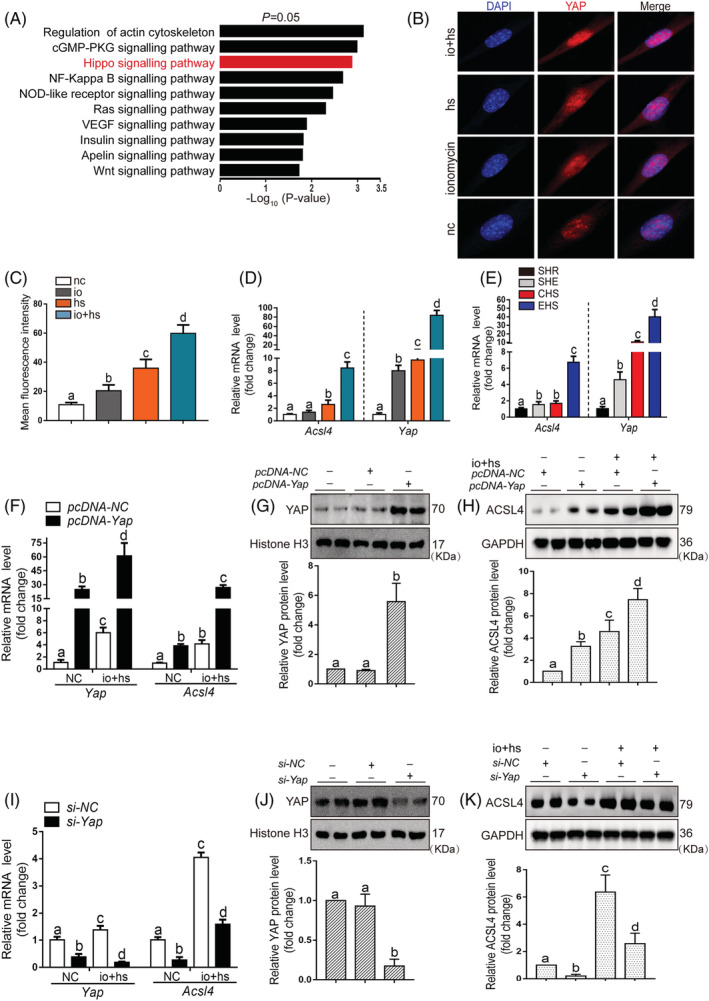
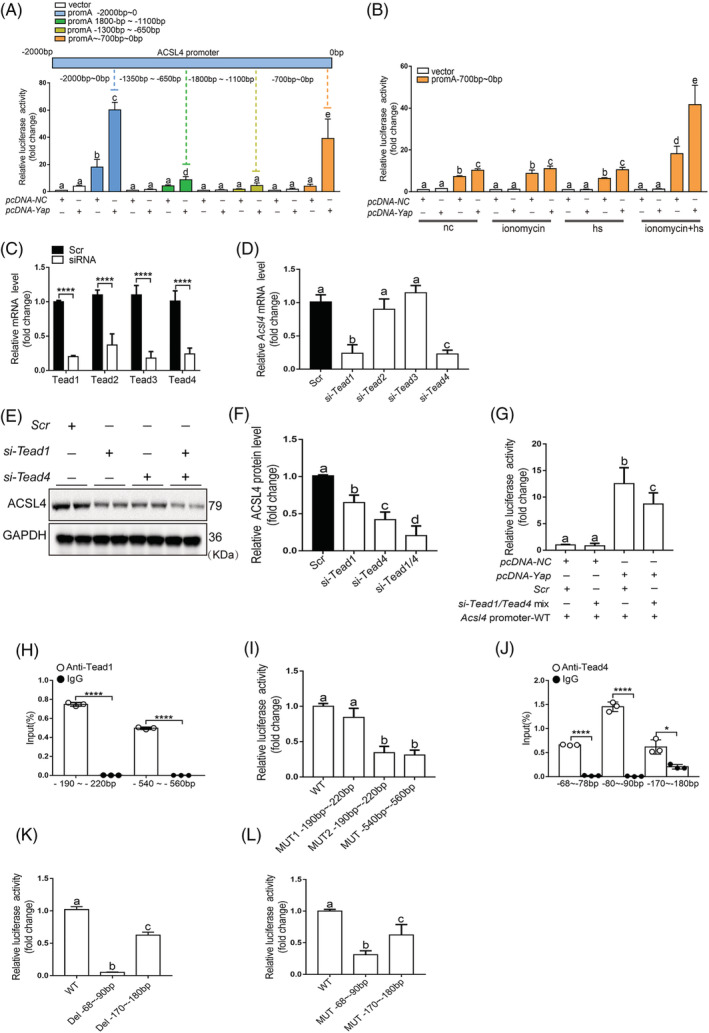
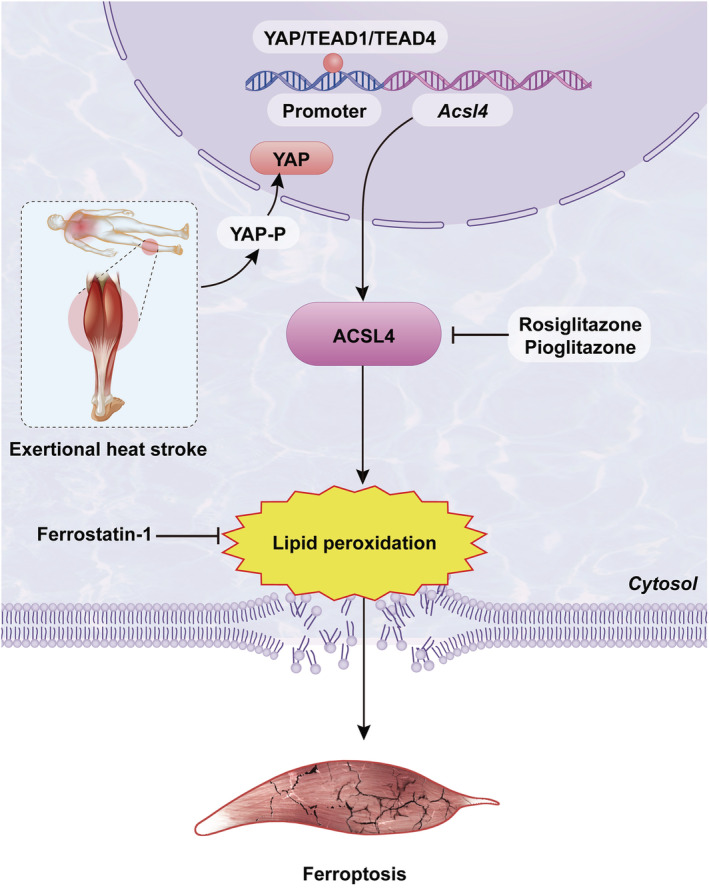
Results: We demonstrated that ferroptosis contributes to RM development following EHS. Ferroptosis inhibitor ferrostatin-1 administration once EHS onset significantly ameliorated the survival rate of EHS mice from 35.357% to 52.288% within 24 h after EHS (P = 0.0028 compared with control) and markedly inhibited RM development induced by EHS. By comparing gene expression of between sham heat rest (SHR) (n = 3) and EHS (n = 3) mice in the gastrocnemius (Gas) muscle tissue, we identified that Acsl4 mRNA expression is elevated in Gas muscle tissue of EHS mice (P = 0.0038 compared with SHR), so as to its protein levels (P = 0.0001 compared with SHR). Followed by increase in creatine kinase (CK) and myoglobin (MB) levels, the labile iron accumulation, decrease in glutathione peroxidase 4 (GPX4) expression, and elevation of lipid peroxidation products. From in vivo and in vitro experiments, inhibition of Acsl4 significantly improves muscle cell death caused by EHS, thereby ameliorating RM development, followed by reduction in CK and MB levels by 30-40% (P < 0.0001; n = 8-10) and 40% (P < 0.0001; n = 8-10), restoration of GPX4 expression, and decrease in lipid peroxidation products. Mechanistically, ACSL4-mediated RM seems to be Yes-associated protein (YAP) dependent via TEA domain transcription factor1/TEA domain transcription factor4.
Conclusions: These findings demonstrate an important role of ACSL4 in mediating ferroptosis activation in the development of RM following EHS and suggest that targeting ACSL4 may represent a novel therapeutic strategy to limit the skeletal muscle cell death and prevent RM after EHS.
Keywords: ACSL4; Exertional heat stroke; Ferroptosis; Lipid peroxidation; Rhabdomyolysis.
© 2022 The Authors. Journal of Cachexia, Sarcopenia and Muscle published by John Wiley & Sons Ltd on behalf of Society on Sarcopenia, Cachexia and Wasting Disorders.
Conflict of interest statement
The authors declare that they have no conflict of interest.
Figures
References
-
- Epstein Y, Yanovich R. Heatstroke. N Engl J Med 2019;380:2449–2459. - PubMed
-
- Laitano O, Oki K, Leon LR. The role of skeletal muscles in exertional heat stroke pathophysiology. Int J Sports Med 2021;42:673–681. - PubMed
-
- Warren JD, Blumbergs PC, Thompson PD. Rhabdomyolysis: a review. Muscle Nerve 2002;25:332–347. - PubMed
-
- Giannoglou GD, Chatzizisis YS, Misirli G. The syndrome of rhabdomyolysis: pathophysiology and diagnosis. Eur J Intern Med 2007;18:90–100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
