

Bile acid-sensitive tuft cells regulate biliary neutrophil influx
- PMID: 35245089
- PMCID: PMC9166270
- DOI: 10.1126/sciimmunol.abj1080
Bile acid-sensitive tuft cells regulate biliary neutrophil influx
Abstract
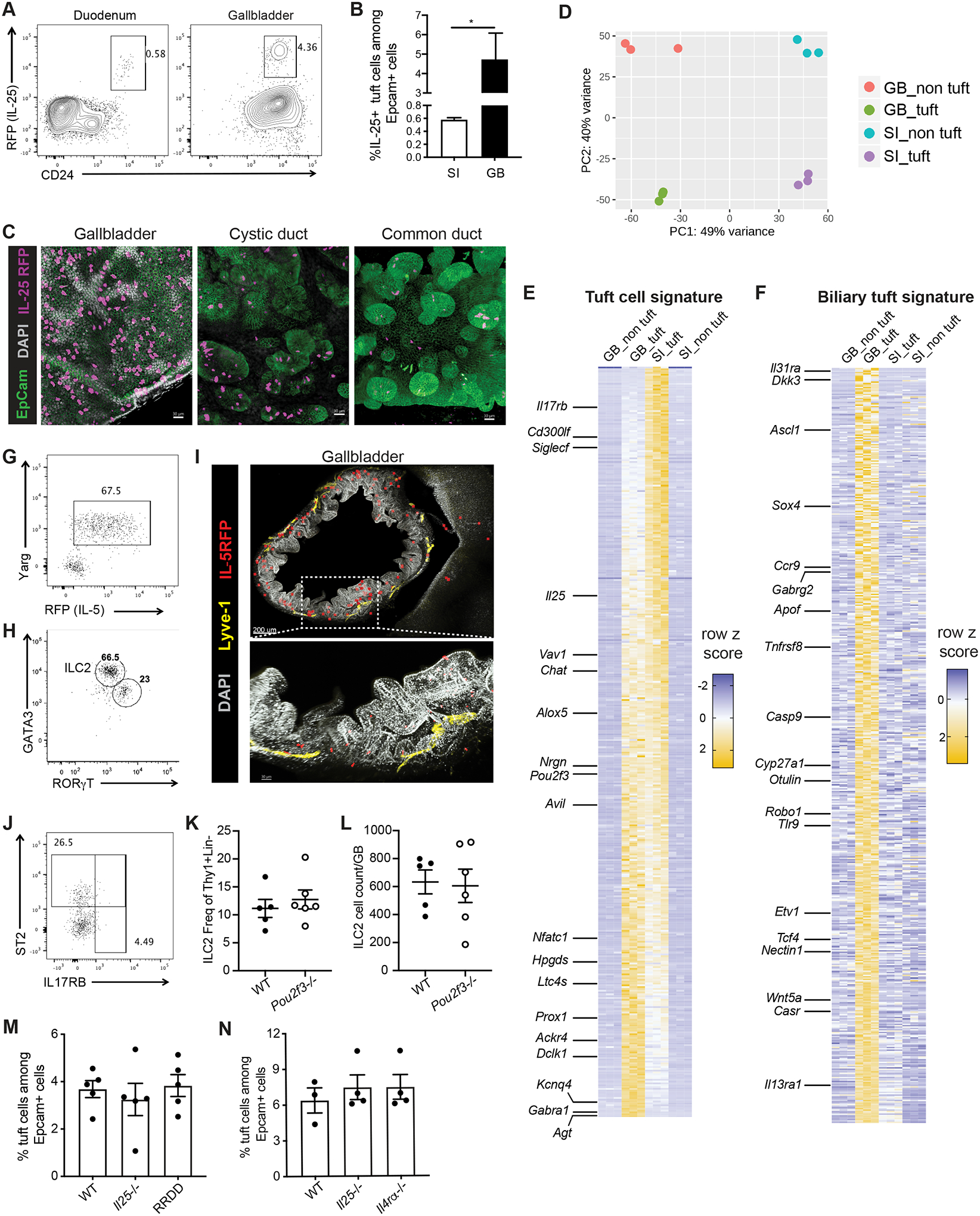
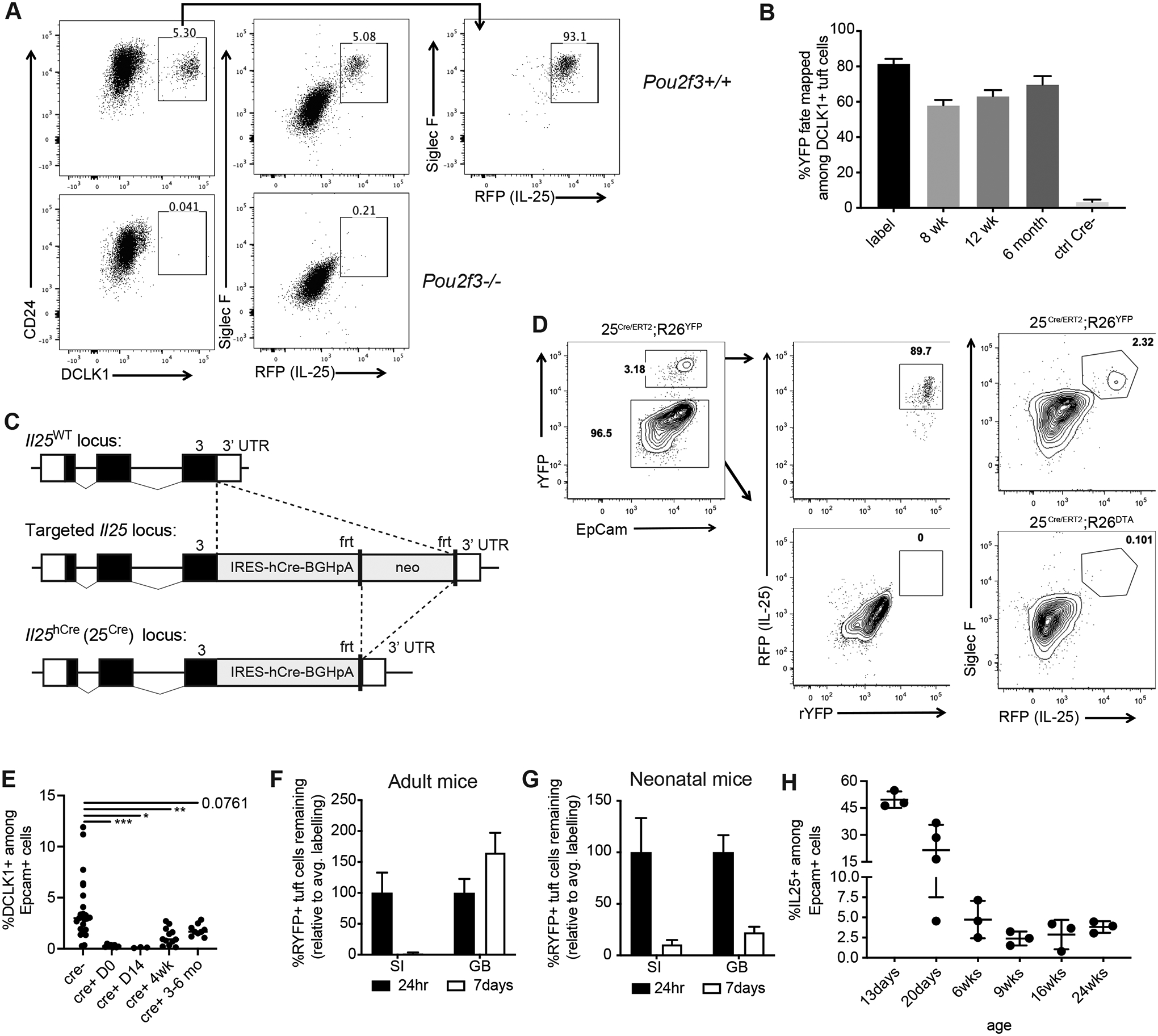
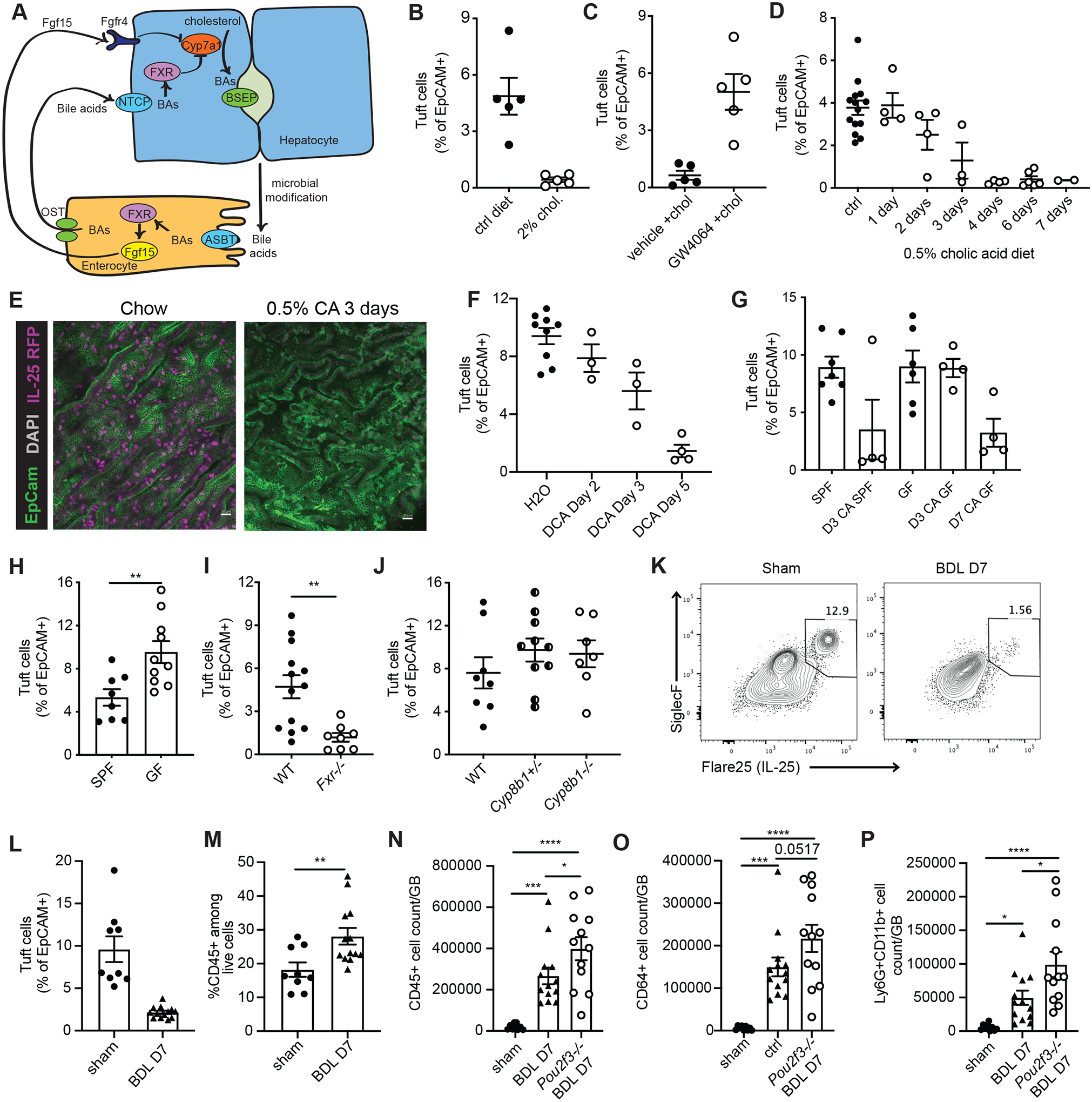
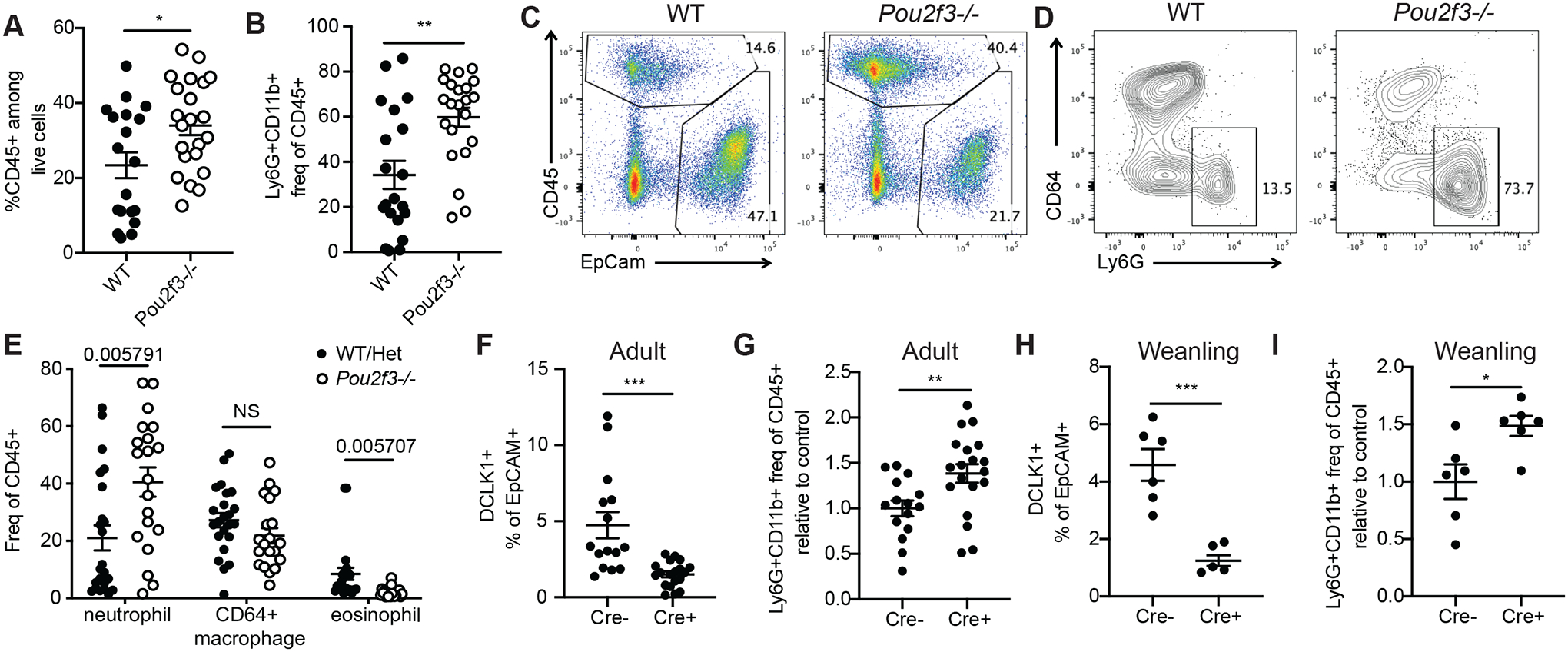
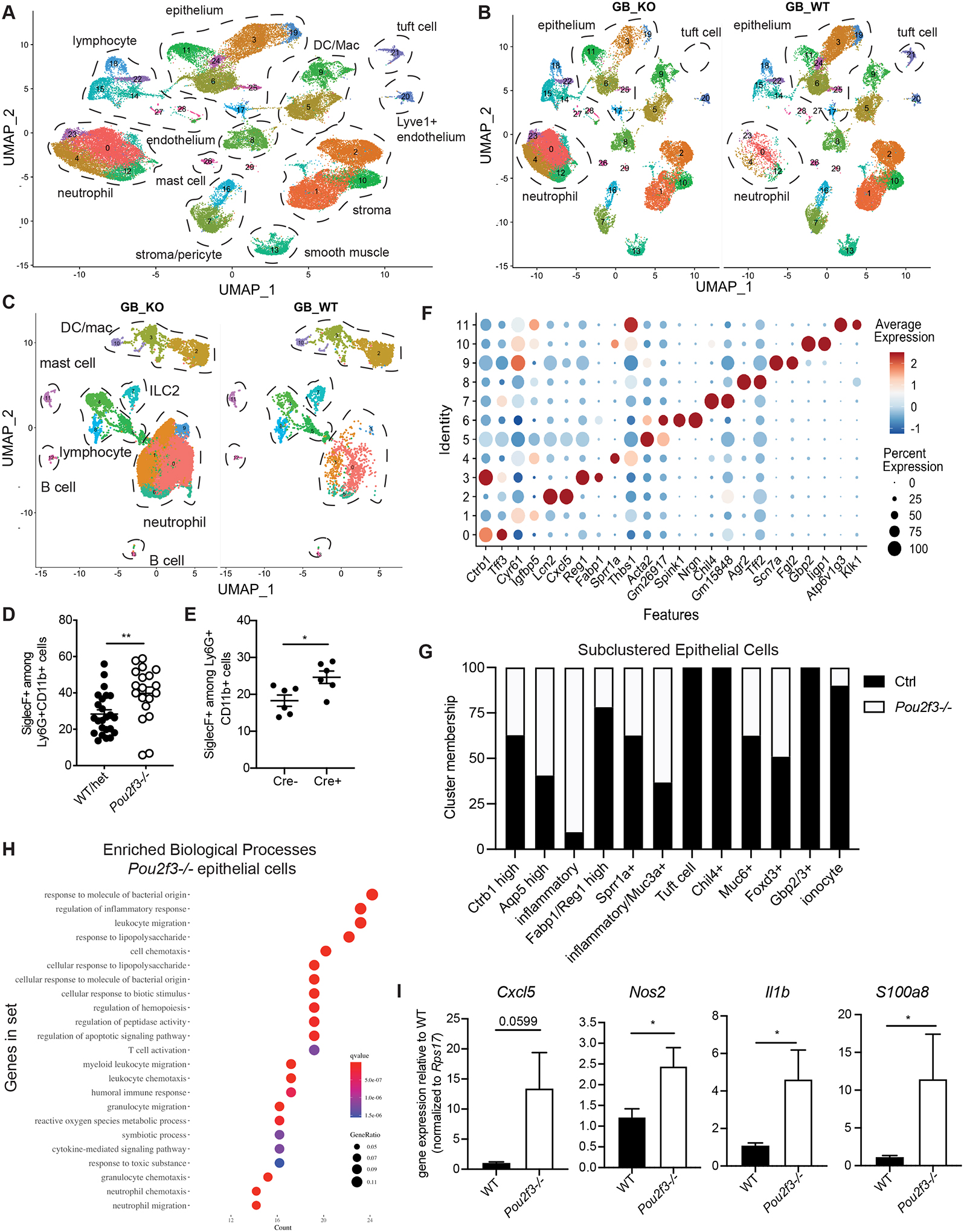
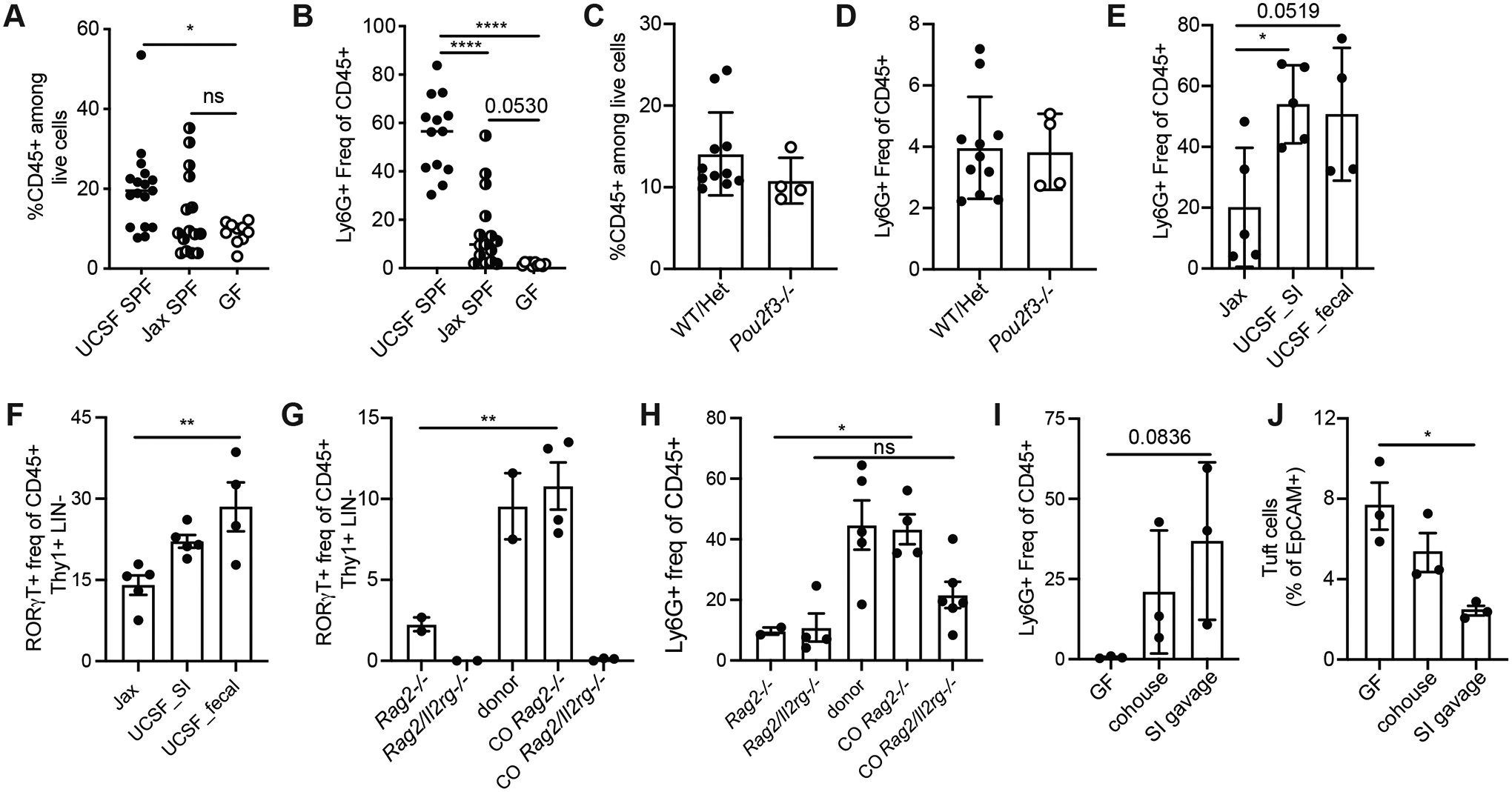
Inflammation and dysfunction of the extrahepatic biliary tree are common causes of human pathology, including gallstones and cholangiocarcinoma. Despite this, we know little about the local regulation of biliary inflammation. Tuft cells, rare sensory epithelial cells, are particularly prevalent in the mucosa of the gallbladder and extrahepatic bile ducts. Here, we show that biliary tuft cells express a core genetic tuft cell program in addition to a tissue-specific gene signature and, in contrast to small intestinal tuft cells, decreased postnatally, coincident with maturation of bile acid production. Manipulation of enterohepatic bile acid recirculation revealed that tuft cell abundance is negatively regulated by bile acids, including in a model of obstructive cholestasis in which inflammatory infiltration of the biliary tree correlated with loss of tuft cells. Unexpectedly, tuft cell-deficient mice spontaneously displayed an increased gallbladder epithelial inflammatory gene signature accompanied by neutrophil infiltration that was modulated by the microbiome. We propose that biliary tuft cells function as bile acid-sensitive negative regulators of inflammation in biliary tissues and serve to limit inflammation under homeostatic conditions.
Conflict of interest statement
Competing interests:
The authors have no competing interests to declare.
Figures
Comment in
-
Bile duct tuft cells regulate immune defences.Nat Rev Immunol. 2022 May;22(5):273. doi: 10.1038/s41577-022-00712-5. Nat Rev Immunol. 2022. PMID: 35318463 No abstract available.
References
-
- Wahlstrom A, Sayin SI, Marschall HU, Backhed F, Intestinal Crosstalk between Bile Acids and Microbiota and Its Impact on Host Metabolism. Cell Metab 24, 41–50 (2016). - PubMed
-
- Gorham F. W. a. I., A. C., General function of the gall bladder from the evolutionary standpoint. American Journal of Digestive Diseases and Nutrition 22, (1938).
-
- Jansen PL, Ghallab A, Vartak N, Reif R, Schaap FG, Hampe J, Hengstler JG, The ascending pathophysiology of cholestatic liver disease. Hepatology (Baltimore, Md.) 65, 722–738 (2017). - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
