

Transitional premonocytes emerge in the periphery for host defense against bacterial infections
- PMID: 35245124
- PMCID: PMC8896792
- DOI: 10.1126/sciadv.abj4641
Transitional premonocytes emerge in the periphery for host defense against bacterial infections
Abstract
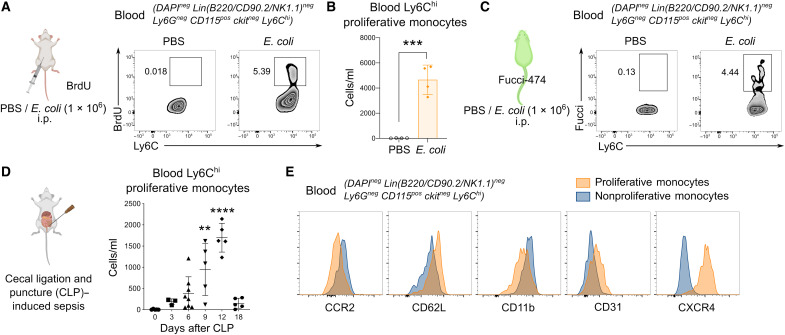
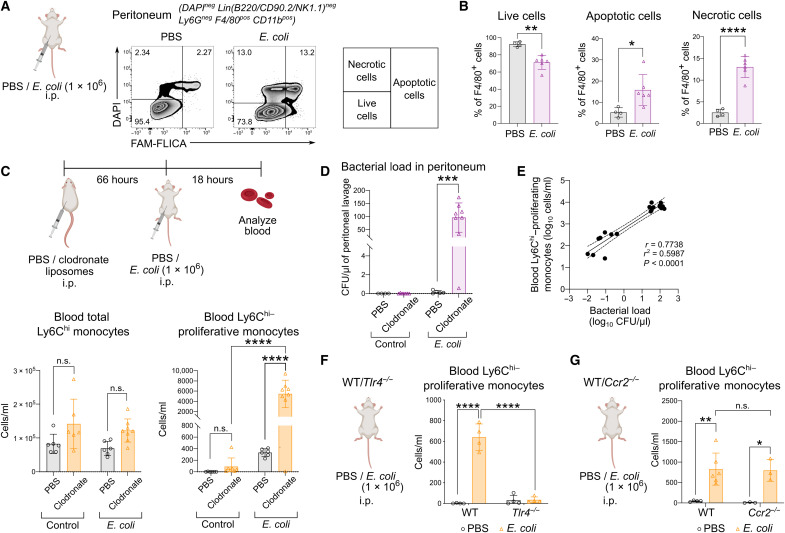
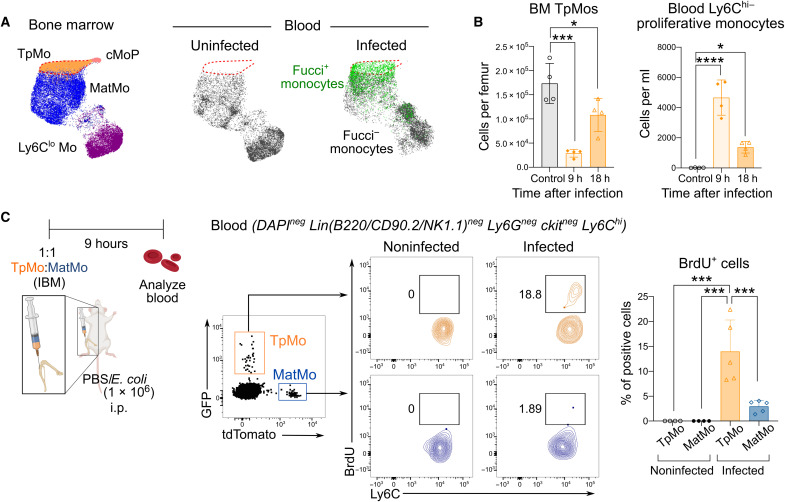
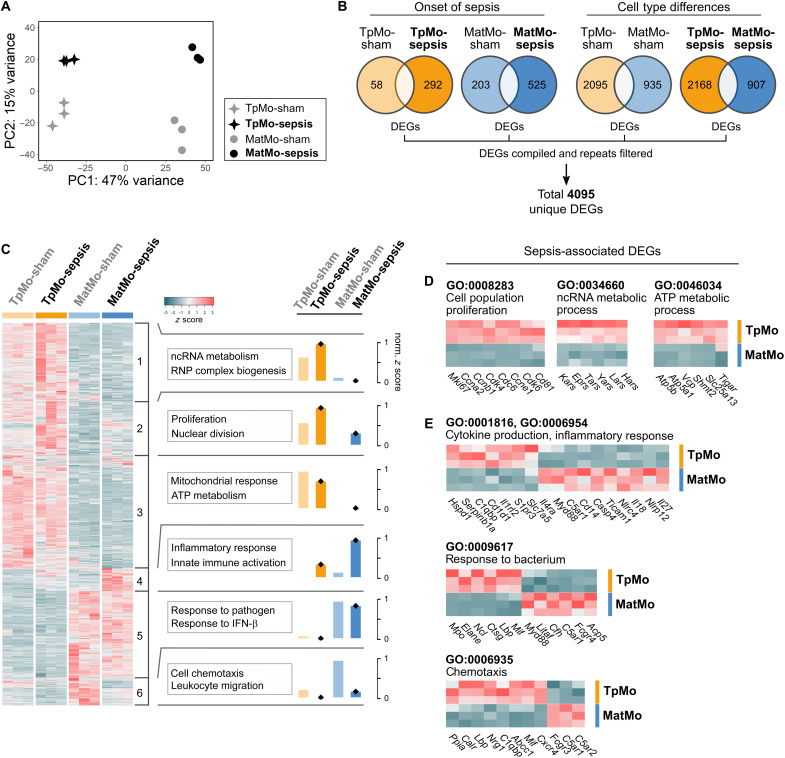
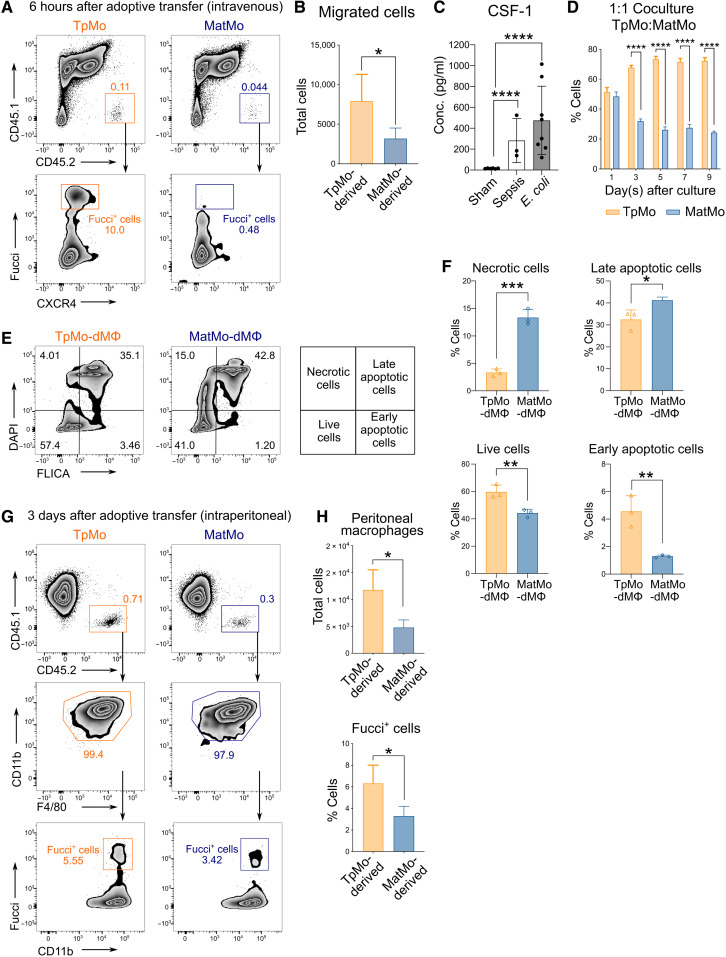
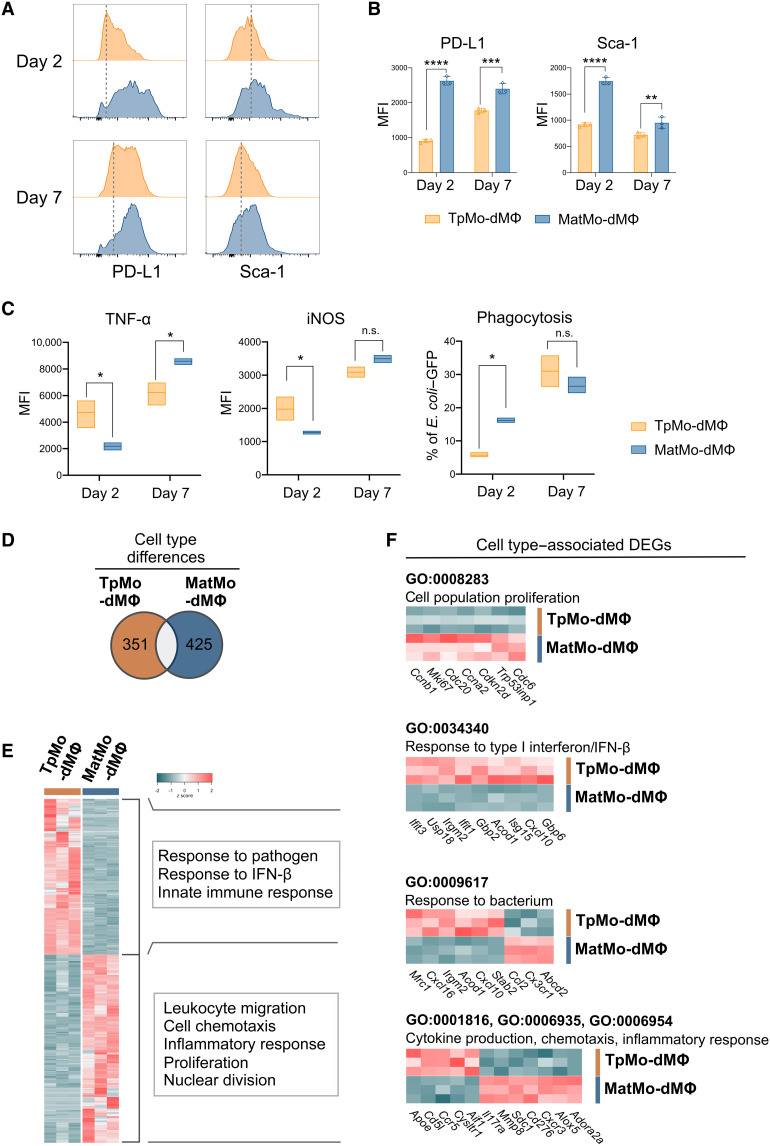
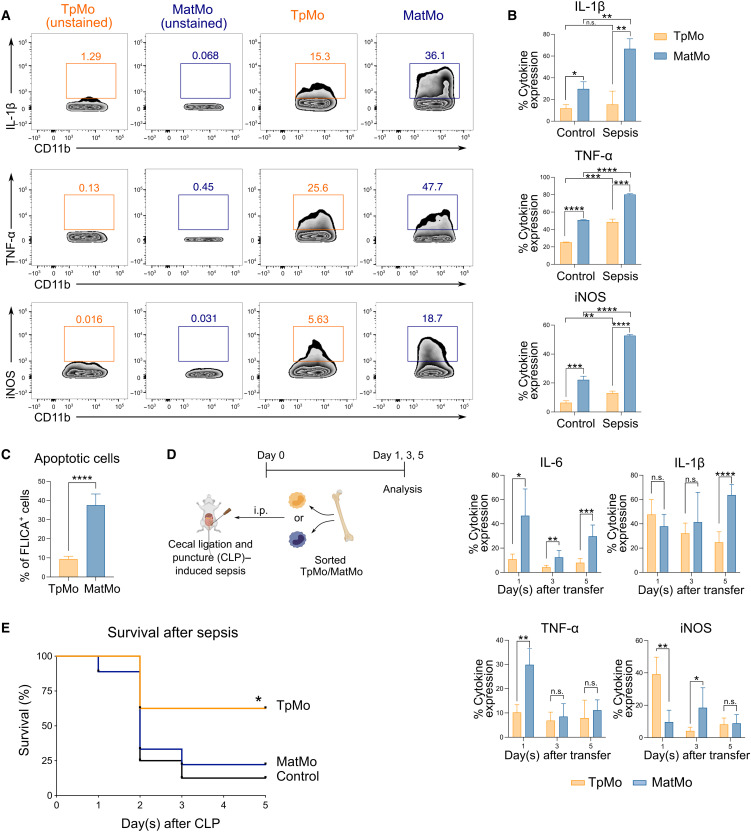
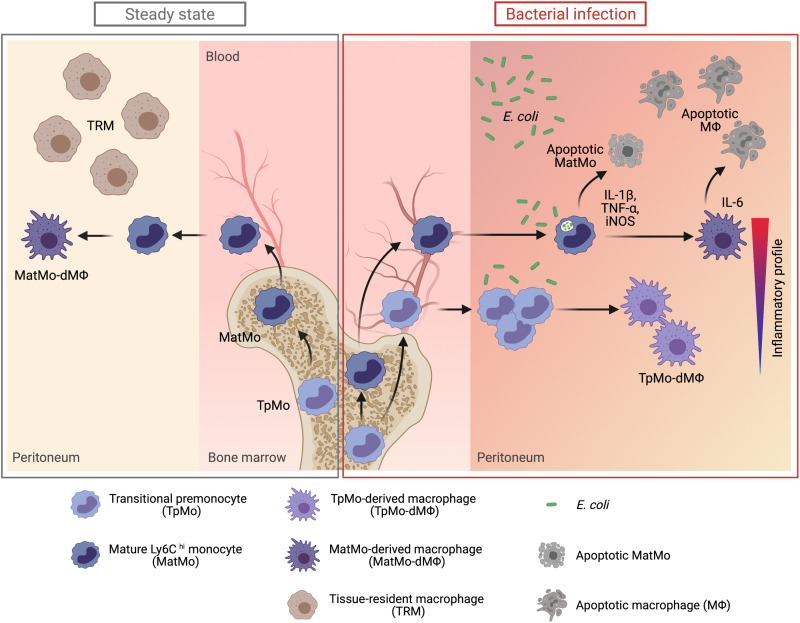
Circulating Ly6Chi monocytes often undergo cellular death upon exhaustion of their antibacterial effector functions, which limits their capacity for subsequent macrophage differentiation. This shrouds the understanding on how the host replaces the tissue-resident macrophage niche effectively during bacterial invasion to avert infection morbidity. Here, we show that proliferating transitional premonocytes (TpMos), an immediate precursor of mature Ly6Chi monocytes (MatMos), were mobilized into the periphery in response to acute bacterial infection and sepsis. TpMos were less susceptible to apoptosis and served as the main source of macrophage replenishment when MatMos were vulnerable toward bacteria-induced cellular death. Furthermore, TpMo and its derived macrophages contributed to host defense by balancing the proinflammatory cytokine response of MatMos. Consequently, adoptive transfer of TpMos improved the survival outcome of lethal sepsis. Our findings hence highlight a protective role for TpMos during bacterial infections and their contribution toward monocyte-derived macrophage heterogeneity in distinct disease outcomes.
Figures
References
-
- Hashimoto D., Chow A., Noizat C., Teo P., Beasley M. B., Leboeuf M., Becker C. D., See P., Price J., Lucas D., Greter M., Mortha A., Boyer S. W., Forsberg E. C., Tanaka M., van Rooijen N., García-Sastre A., Stanley E. R., Ginhoux F., Frenette P. S., Merad M., Tissue-resident macrophages self-maintain locally throughout adult life with minimal contribution from circulating monocytes. Immunity 38, 792–804 (2013). - PMC - PubMed
-
- Ginhoux F., Guilliams M., Tissue-resident macrophage ontogeny and homeostasis. Immunity. 44, 439–449 (2016). - PubMed
-
- Zhang N., Czepielewski R. S., Jarjour N. N., Erlich E. C., Esaulova E., Saunders B. T., Grover S. P., Cleuren A. C., Broze G. J., Edelson B. T., Mackman N., Zinselmeyer B. H., Randolph G. J., Expression of factor V by resident macrophages boosts host defense in the peritoneal cavity. J. Exp. Med. 216, 1291–1300 (2019). - PMC - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
