

The molecular basis of FimT-mediated DNA uptake during bacterial natural transformation
- PMID: 35246533
- PMCID: PMC8897410
- DOI: 10.1038/s41467-022-28690-1
The molecular basis of FimT-mediated DNA uptake during bacterial natural transformation
Abstract
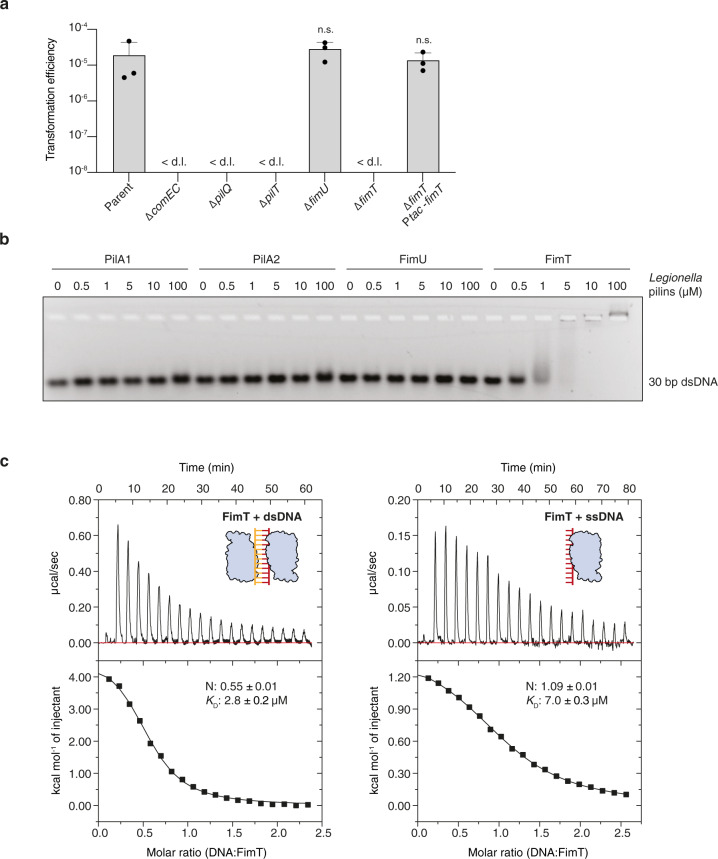
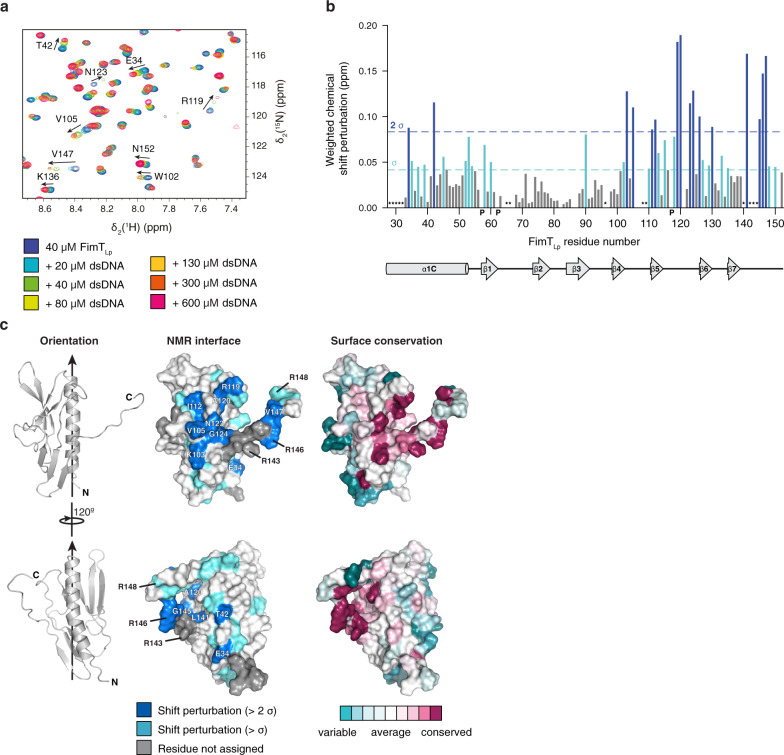
Naturally competent bacteria encode sophisticated protein machinery for the uptake and translocation of exogenous DNA into the cell. If this DNA is integrated into the bacterial genome, the bacterium is said to be naturally transformed. Most competent bacterial species utilise type IV pili for the initial DNA uptake step. These proteinaceous cell-surface structures are composed of thousands of pilus subunits (pilins), designated as major or minor according to their relative abundance in the pilus. Here, we show that the minor pilin FimT plays an important role in the natural transformation of Legionella pneumophila. We use NMR spectroscopy, in vitro DNA binding assays and in vivo transformation assays to understand the molecular basis of FimT's role in this process. FimT binds to DNA via an electropositive patch, rich in arginines, several of which are well-conserved and located in a conformationally flexible C-terminal tail. FimT orthologues from other Gammaproteobacteria share the ability to bind to DNA. Our results suggest that FimT plays an important role in DNA uptake in a wide range of competent species.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Johnsborg O, Eldholm V, Håvarstein LS. Natural genetic transformation: prevalence, mechanisms and function. Res. Microbiol. 2007;158:767–778. - PubMed
-
- Johnston C, Martin B, Fichant G, Polard P, Claverys J-P. Bacterial transformation: distribution, shared mechanisms and divergent control. Nat. Rev. Microbiol. 2014;12:181–196. - PubMed
-
- Sánchez-Busó L, Comas I, Jorques G, González-Candelas F. Recombination drives genome evolution in outbreak-related Legionella pneumophila isolates. Nat. Genet. 2014;46:1205–1211. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
