

Different responses to DNA damage determine ageing differences between organs
- PMID: 35246937
- PMCID: PMC9009128
- DOI: 10.1111/acel.13562
Different responses to DNA damage determine ageing differences between organs
Abstract
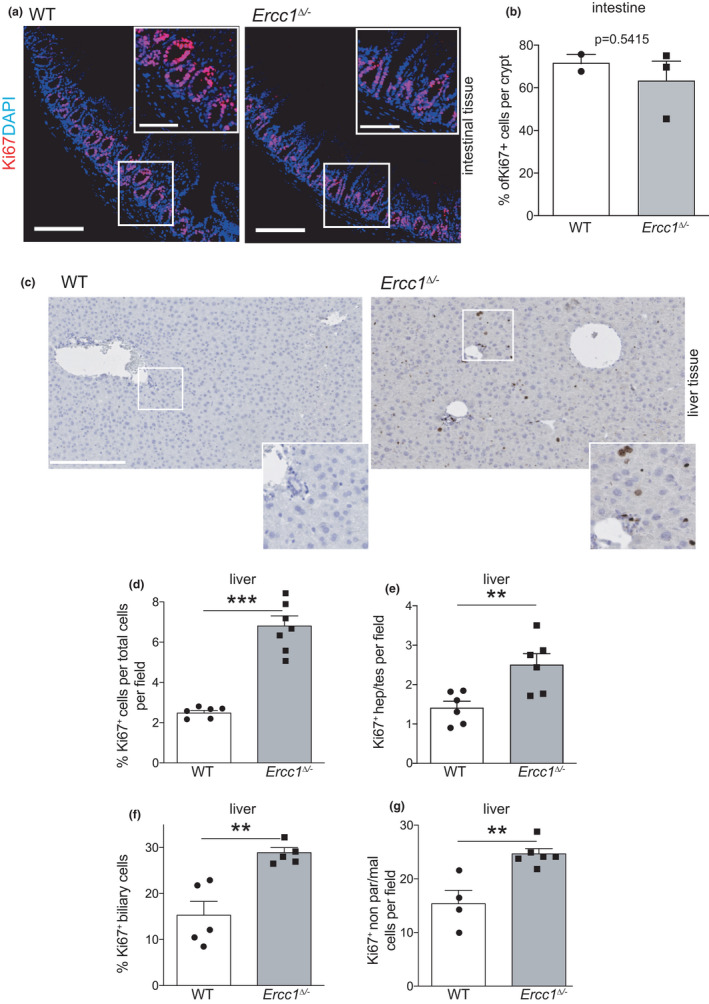
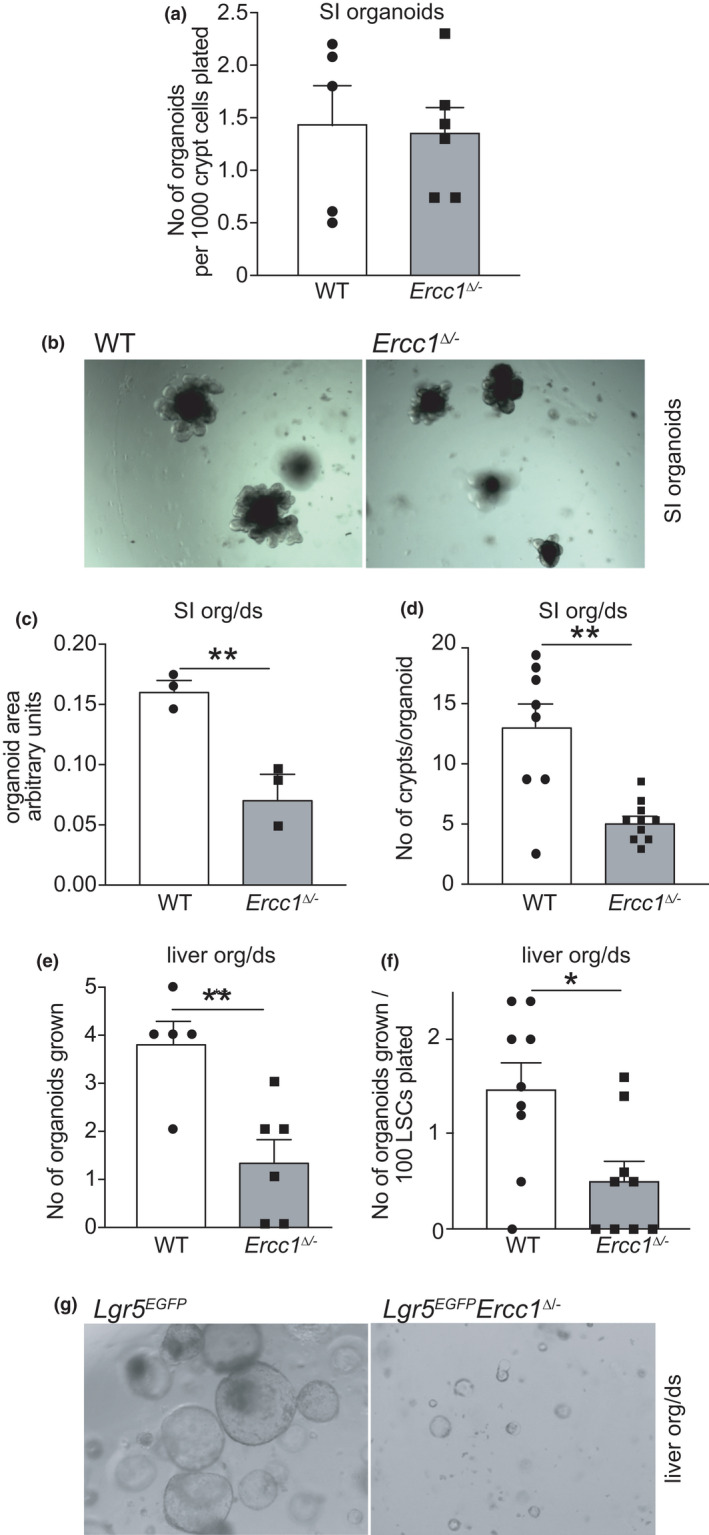
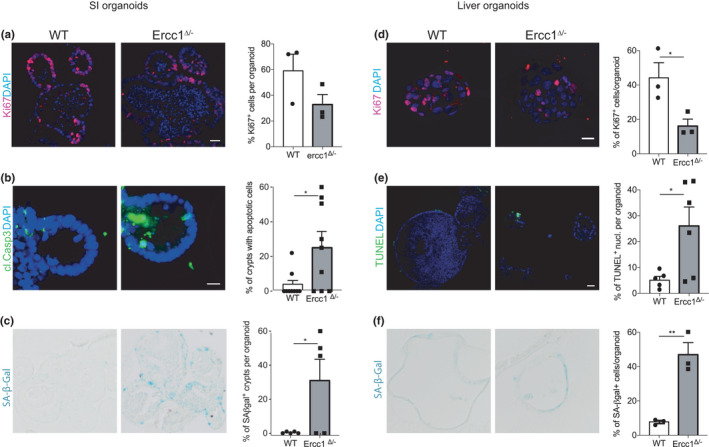
Organs age differently, causing wide heterogeneity in multimorbidity, but underlying mechanisms are largely elusive. To investigate the basis of organ-specific ageing, we utilized progeroid repair-deficient Ercc1Δ/- mouse mutants and systematically compared at the tissue, stem cell and organoid level two organs representing ageing extremes. Ercc1Δ/- intestine shows hardly any accelerated ageing. Nevertheless, we found apoptosis and reduced numbers of intestinal stem cells (ISCs), but cell loss appears compensated by over-proliferation. ISCs retain their organoid-forming capacity, but organoids perform poorly in culture, compared with WT. Conversely, liver ages dramatically, even causing early death in Ercc1-KO mice. Apoptosis, p21, polyploidization and proliferation of various (stem) cells were prominently elevated in Ercc1Δ/- liver and stem cell populations were either largely unaffected (Sox9+), or expanding (Lgr5+), but were functionally exhausted in organoid formation and development in vitro. Paradoxically, while intestine displays less ageing, repair in WT ISCs appears inferior to liver as shown by enhanced sensitivity to various DNA-damaging agents, and lower lesion removal. Our findings reveal organ-specific anti-ageing strategies. Intestine, with short lifespan limiting time for damage accumulation and repair, favours apoptosis of damaged cells relying on ISC plasticity. Liver with low renewal rates depends more on repair pathways specifically protecting the transcribed compartment of the genome to promote sustained functionality and cell preservation. As shown before, the hematopoietic system with intermediate self-renewal mainly invokes replication-linked mechanisms, apoptosis and senescence. Hence, organs employ different genome maintenance strategies, explaining heterogeneity in organ ageing and the segmental nature of DNA-repair-deficient progerias.
Keywords: DNA damage response; ERCC1; adult stem cells; genome maintenance; liver; nucleotide excision repair; organoids; small intestine.
© 2022 The Authors. Aging Cell published by the Anatomical Society and John Wiley & Sons Ltd.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Ahmad, A. , Robinson, A. R. , Duensing, A. , van Drunen, E. , Beverloo, H. B. , Weisberg, D. B. , Hasty, P. , Hoeijmakers, J. H. , & Niedernhofer, L. J. (2008). ERCC1‐XPF endonuclease facilitates DNA double‐strand break repair. Molecular and Cellular Biology, 28, 5082–5092. 10.1128/MCB.00293-08 - DOI - PMC - PubMed
-
- Alyodawi, K. , Vermeij, W. P. , Omairi, S. , Kretz, O. , Hopkinson, M. , Solagna, F. , Joch, B. , Brandt, R. M. C. , Barnhoorn, S. , Vliet, N. , Ridwan, Y. , Essers, J. , Mitchell, R. , Morash, T. , Pasternack, A. , Ritvos, O. , Matsakas, A. , Collins‐Hooper, H. , Huber, T. B. , … Patel, K. (2019). Compression of morbidity in a progeroid mouse model through the attenuation of myostatin/activin signalling. Journal of Cachexia, Sarcopenia and Muscle, 10, 662–686. 10.1002/jcsm.12404 - DOI - PMC - PubMed
-
- Andressoo, J.‐O. , Mitchell, J. R. , de Wit, J. , Hoogstraten, D. , Volker, M. , Toussaint, W. , Speksnijder, E. , Beems, R. B. , van Steeg, H. , Jans, J. , de Zeeuw, C. I. , Jaspers, N. G. J. , Raams, A. , Lehmann, A. R. , Vermeulen, W. , Hoeijmakers, J. H. J. , & van der Horst, G. T. J. (2006). An Xpd mouse model for the combined xeroderma pigmentosum/Cockayne syndrome exhibiting both cancer predisposition and segmental progeria. Cancer Cell, 10, 121–132. 10.1016/j.ccr.2006.05.027 - DOI - PubMed
-
- Baar, M. P. , Brandt, R. M. C. , Putavet, D. A. , Klein, J. D. D. , Derks, K. W. J. , Bourgeois, B. R. M. , Stryeck, S. , Rijksen, Y. , van Willigenburg, H. , Feijtel, D. A. , van der Pluijm, I. , Essers, J. , van Cappellen, W. A. , van IJcken, W. F. , Houtsmuller, A. B. , Pothof, J. , de Bruin, R. W. F. , Madl, T. , Hoeijmakers, J. H. J. , … de Keizer, P. L. J. (2017). Targeted apoptosis of senescent cells restores tissue homeostasis in response to chemotoxicity and aging. Cell, 169(132–147), e116. 10.1016/j.cell.2017.02.031 - DOI - PMC - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
