

Exercise training remodels subcutaneous adipose tissue in adults with obesity even without weight loss
- PMID: 35249225
- PMCID: PMC9058215
- DOI: 10.1113/JP282371
Exercise training remodels subcutaneous adipose tissue in adults with obesity even without weight loss
Abstract
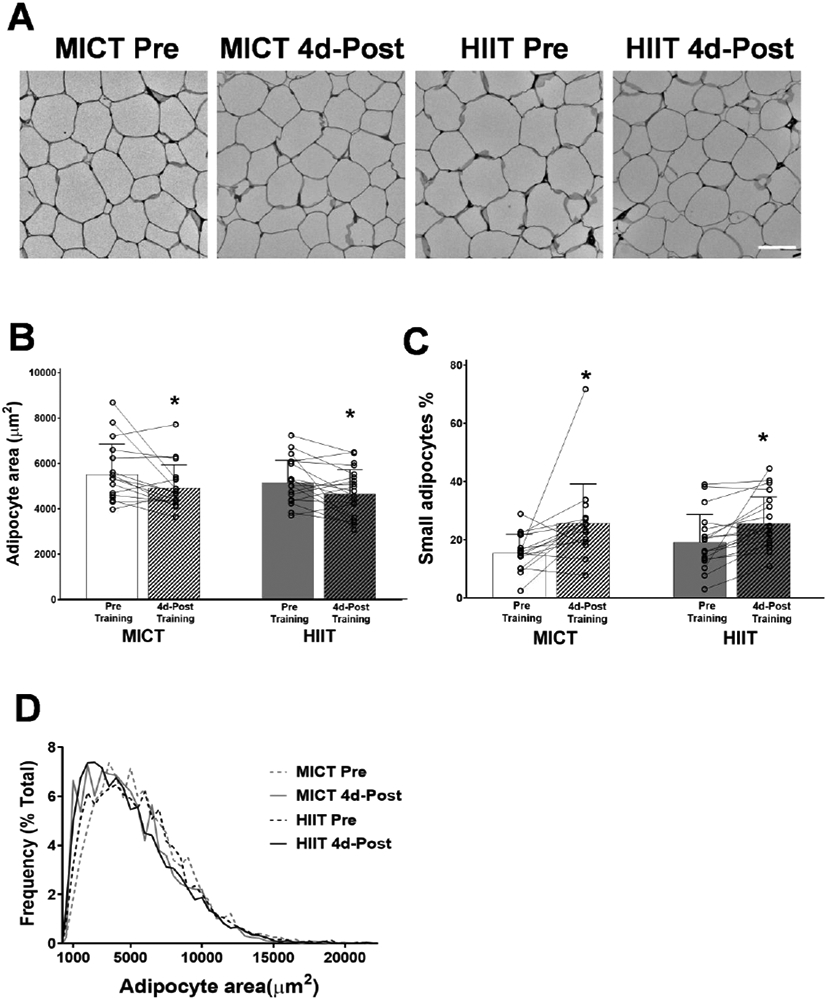
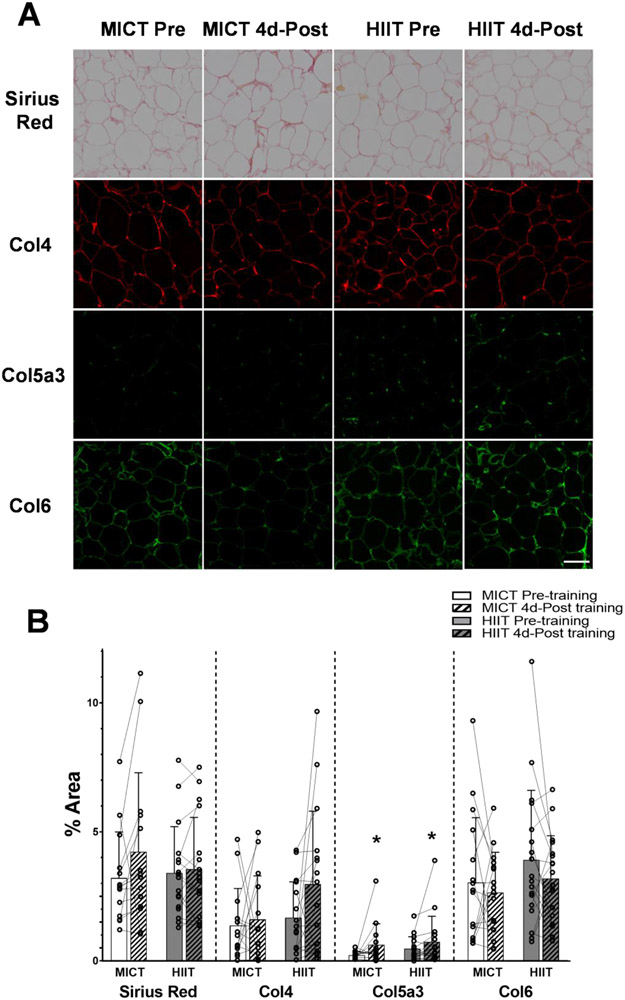
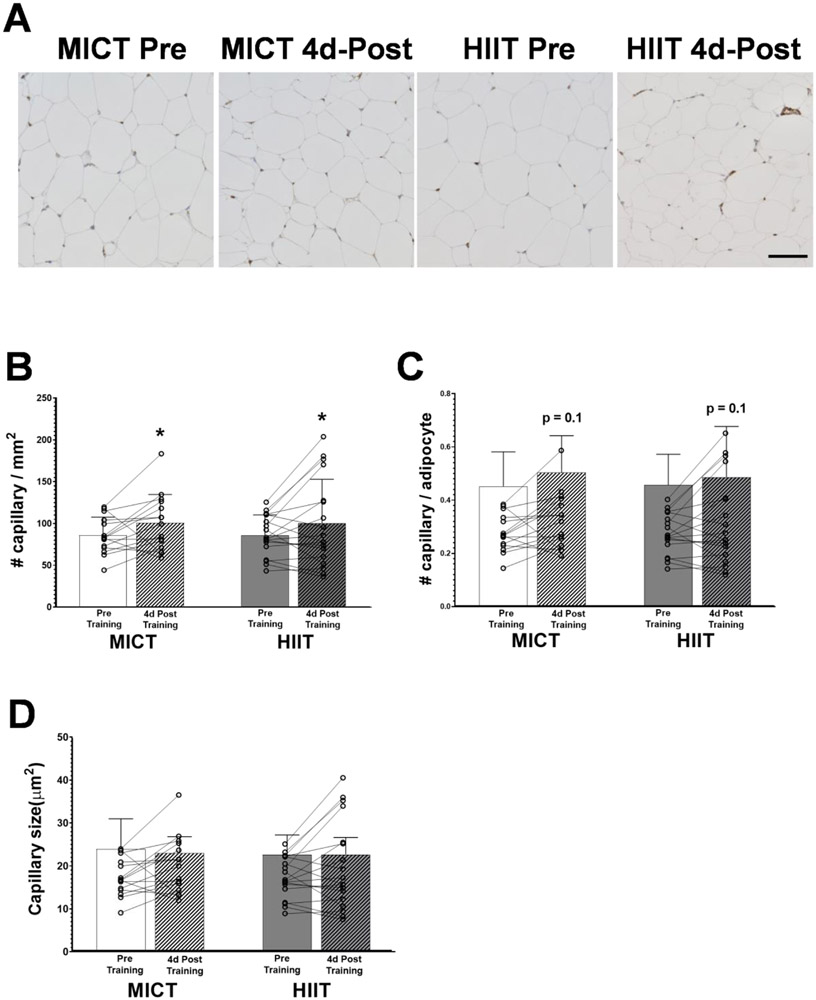
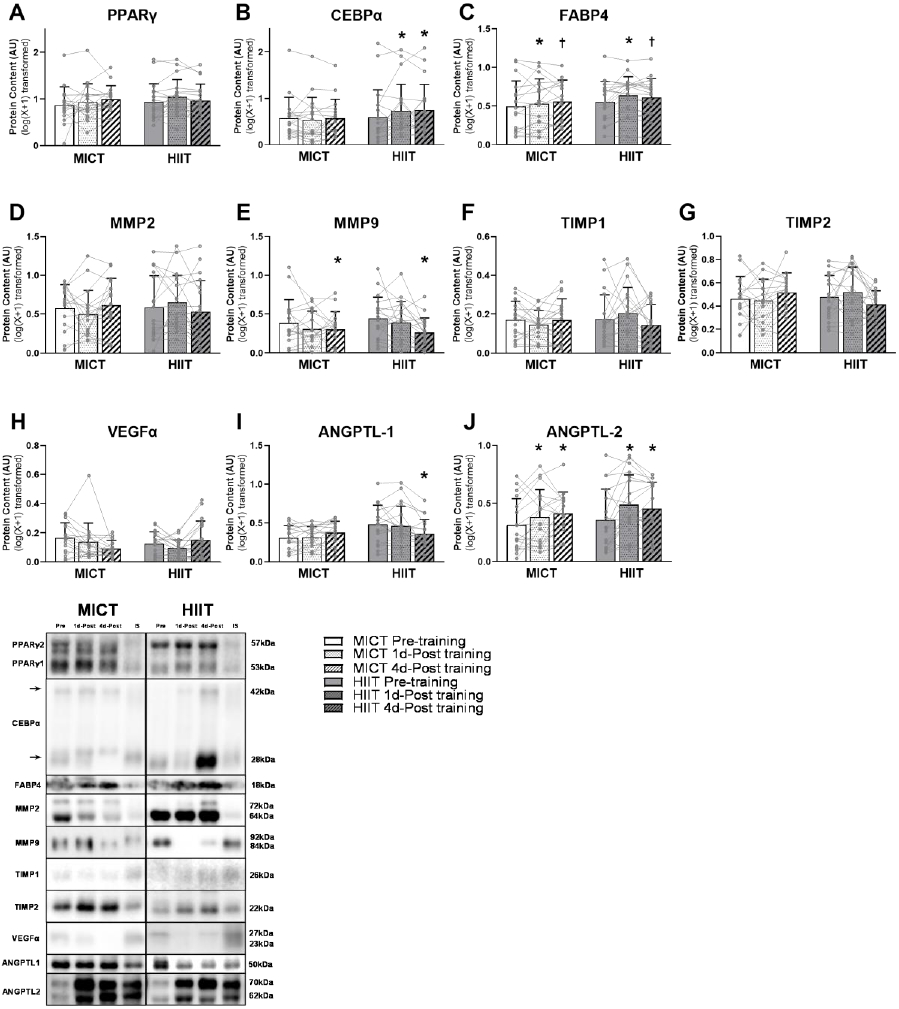
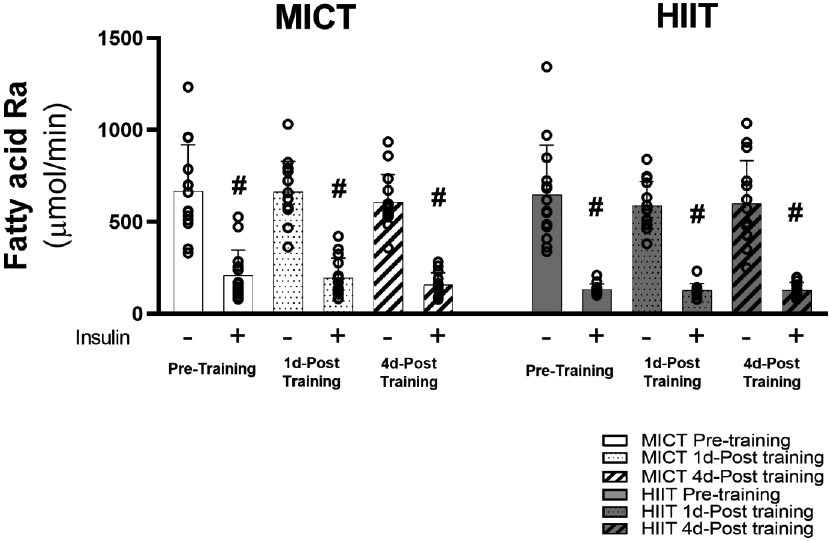
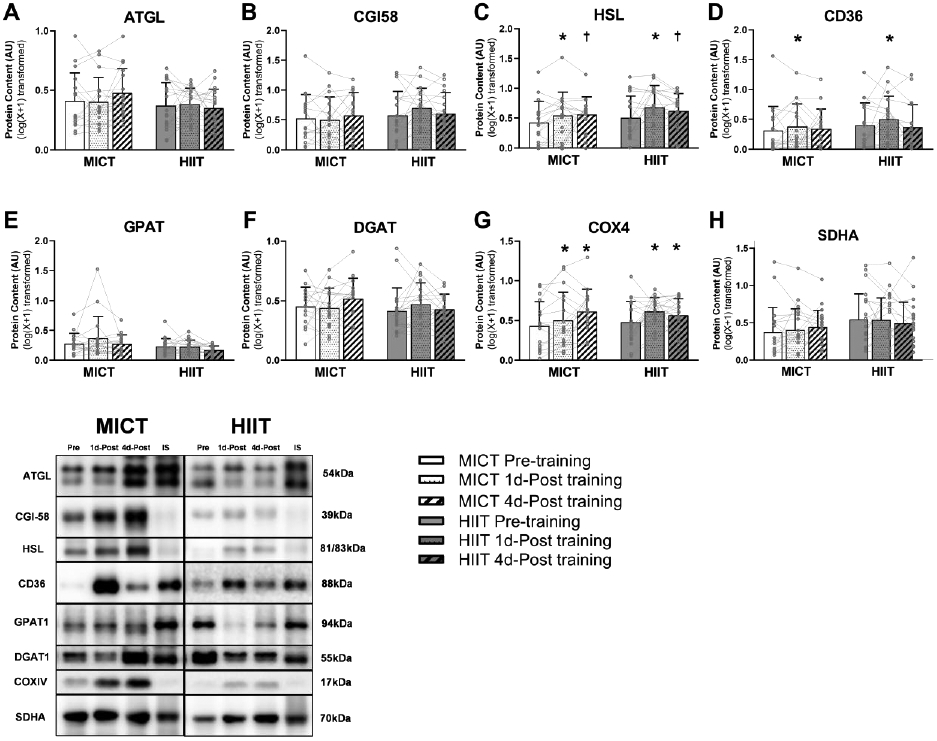
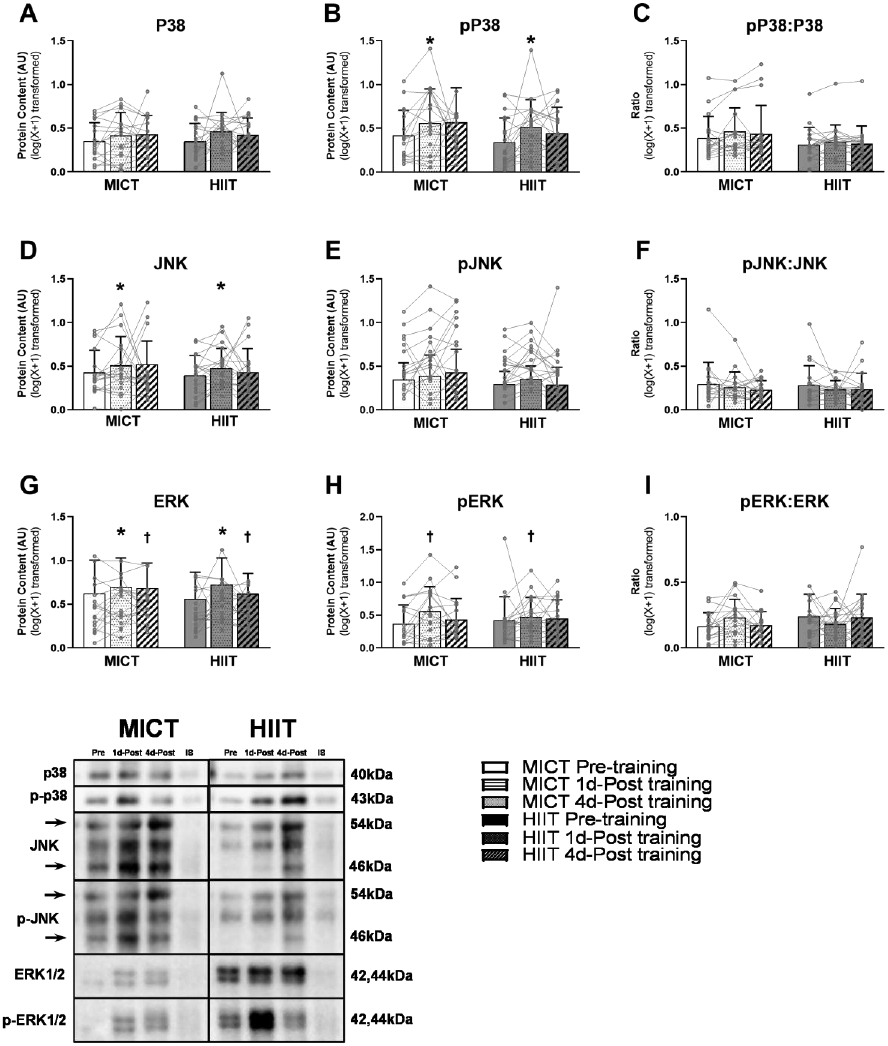
Excessive adipose tissue mass underlies much of the metabolic health complications in obesity. Although exercise training is known to improve metabolic health in individuals with obesity, the effects of exercise training without weight loss on adipose tissue structure and metabolic function remain unclear. Thirty-six adults with obesity (body mass index = 33 ± 3 kg · m-2 ) were assigned to 12 weeks (4 days week-1 ) of either moderate-intensity continuous training (MICT; 70% maximal heart rate, 45 min; n = 17) or high-intensity interval training (HIIT; 90% maximal heart rate, 10 × 1 min; n = 19), maintaining their body weight throughout. Abdominal subcutaneous adipose tissue (aSAT) biopsy samples were collected once before and twice after training (1 day after last exercise and again 4 days later). Exercise training modified aSAT morphology (i.e. reduced fat cell size, increased collagen type 5a3, both P ≤ 0.05, increased capillary density, P = 0.05) and altered protein abundance of factors that regulate aSAT remodelling (i.e. reduced matrix metallopeptidase 9; P = 0.02; increased angiopoietin-2; P < 0.01). Exercise training also increased protein abundance of factors that regulate lipid metabolism (e.g. hormone sensitive lipase and fatty acid translocase; P ≤ 0.03) and key proteins involved in the mitogen-activated protein kinase pathway when measured the day after the last exercise session. However, most of these exercise-mediated changes were no longer significant 4 days after exercise. Importantly, MICT and HIIT induced remarkably similar adaptations in aSAT. Collectively, even in the absence of weight loss, 12 weeks of exercise training induced changes in aSAT structure, as well as factors that regulate metabolism and the inflammatory signal pathway in adults with obesity. KEY POINTS: Exercise training is well-known to improve metabolic health in obesity, although how exercise modifies the structure and metabolic function of adipose tissue, in the absence of weight loss, remains unclear. We report that both 12 weeks of moderate-intensity continuous training (MICT) and 12 weeks of high-intensity interval training (HIIT) induced modifications in adipose tissue structure and factors that regulate adipose tissue remodelling, metabolism and the inflammatory signal pathway in adults with obesity, even without weight loss (with no meaningful differences between MICT and HIIT). The modest modifications in adipose tissue structure in response to 12 weeks of MICT or HIIT did not lead to changes in the rate of fatty acid release from adipose tissue. These results expand our understanding about the effects of two commonly used exercise training prescriptions (MICT and HIIT) on adipose tissue remodelling that may lead to advanced strategies for improving metabolic health outcomes in adults with obesity.
Keywords: adipose tissue; exercise training; high-intensity interval training.
© 2022 The Authors. The Journal of Physiology © 2022 The Physiological Society.
Figures

Comment in
-
Favourable alterations in adipose remodelling induced by exercise training without weight loss: exploring the role of microvascular endothelial function.J Physiol. 2022 Aug;600(16):3647-3650. doi: 10.1113/JP283091. Epub 2022 Jul 21. J Physiol. 2022. PMID: 35833700 Free PMC article. No abstract available.
References
-
- Camp HS & Tafuri SR. (1997). Regulation of Peroxisome Proliferator-activated Receptor γ Activity by Mitogen-activated Protein Kinase*. Journal of Biological Chemistry 272, 10811–10816. - PubMed
-
- Chen X-H, Zhao Y-P, Xue M, Ji C-B, Gao C-L, Zhu J-G, Qin D-N, Kou C-Z, Qin X-H & Tong M-L. (2010). TNF-α induces mitochondrial dysfunction in 3T3-L1 adipocytes. Molecular and cellular endocrinology 328, 63–69. - PubMed