Timing of readiness potentials reflect a decision-making process in the human brain
- PMID: 35252759
- PMCID: PMC8896820
- DOI: 10.1007/s42113-020-00097-5
Timing of readiness potentials reflect a decision-making process in the human brain
Abstract
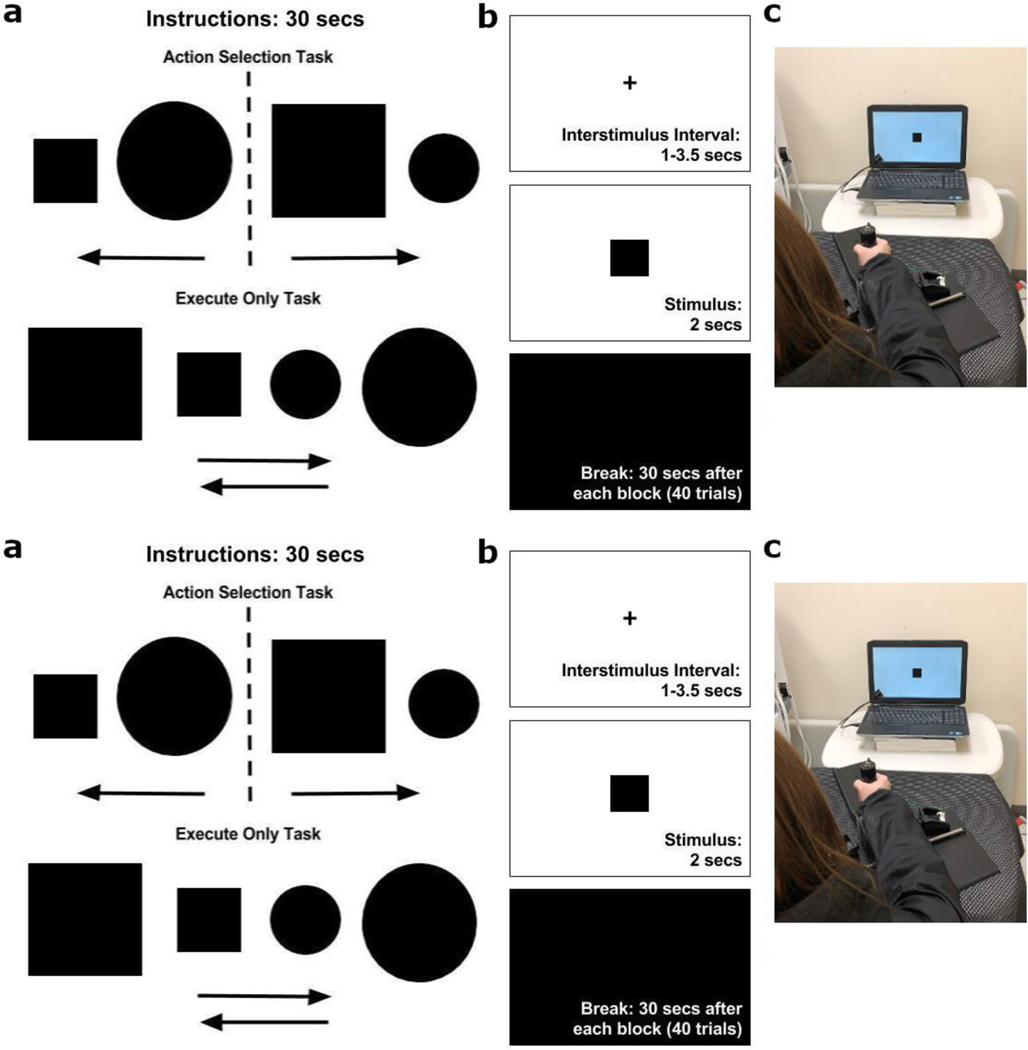
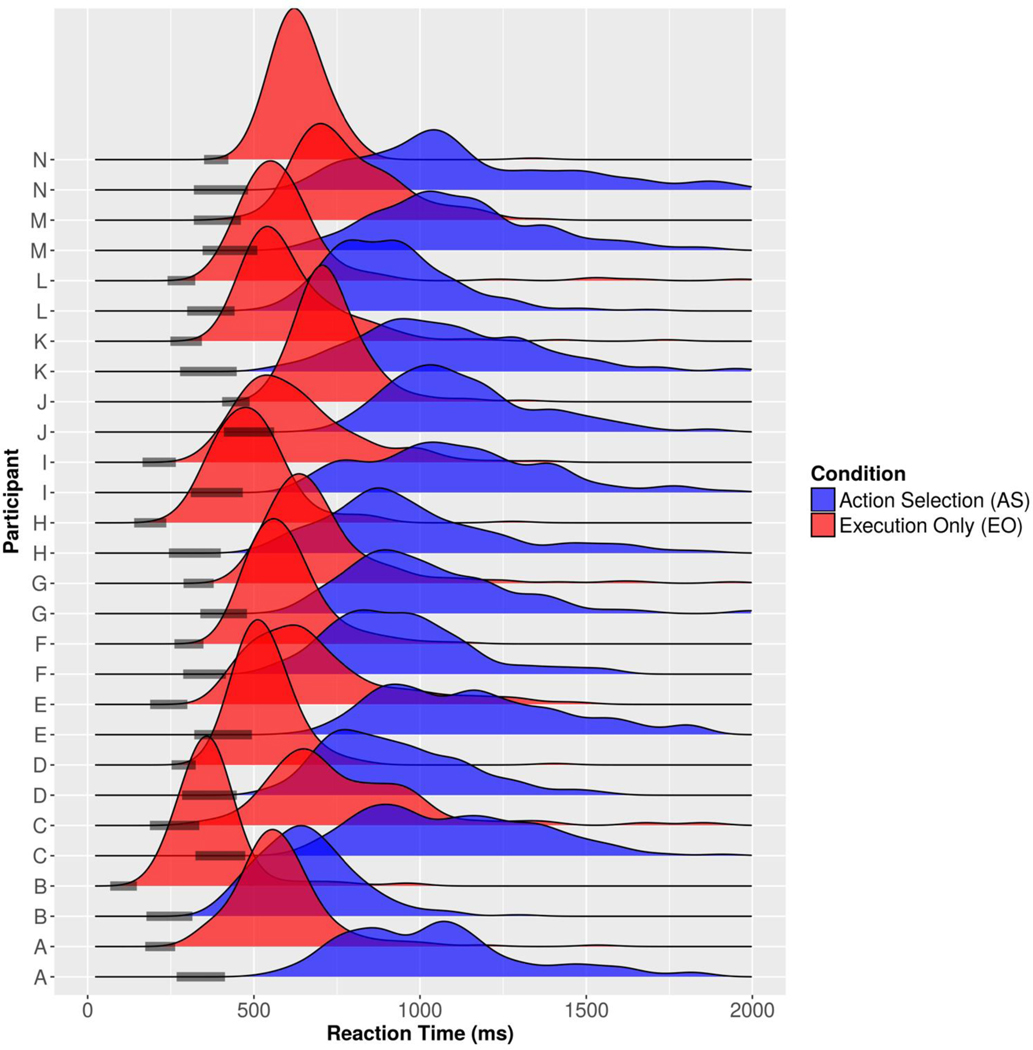
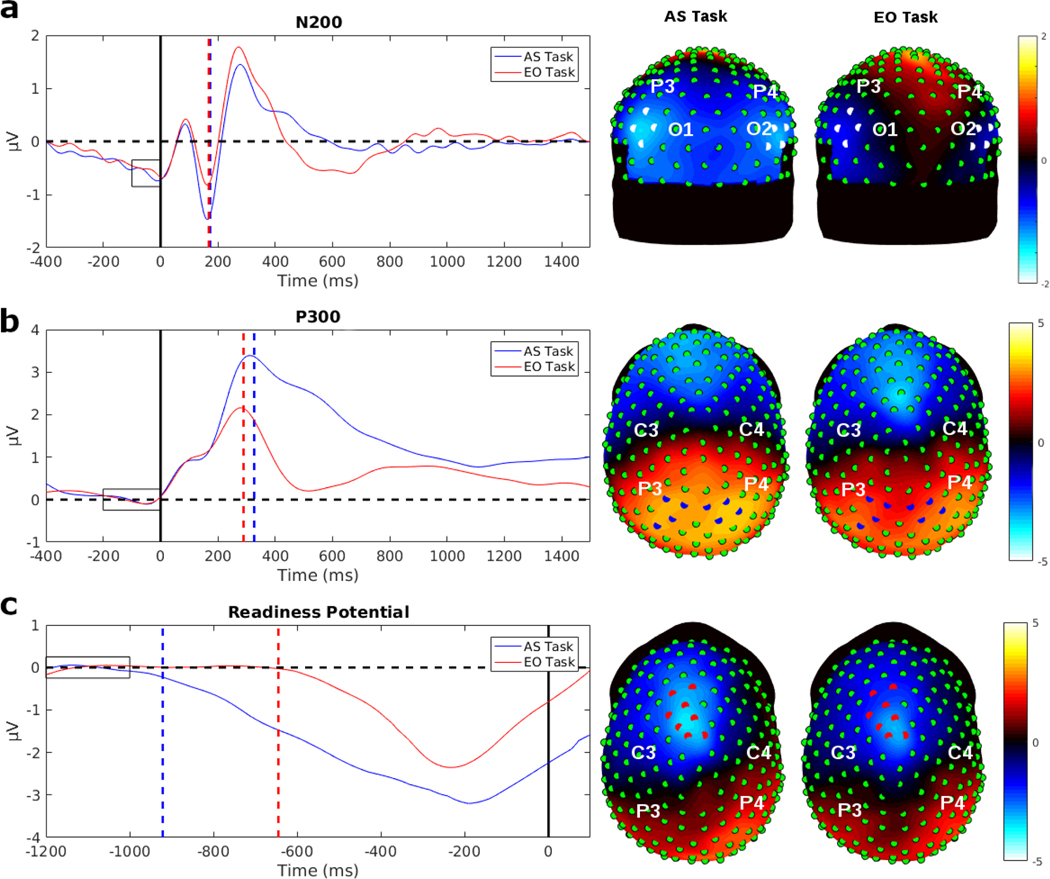
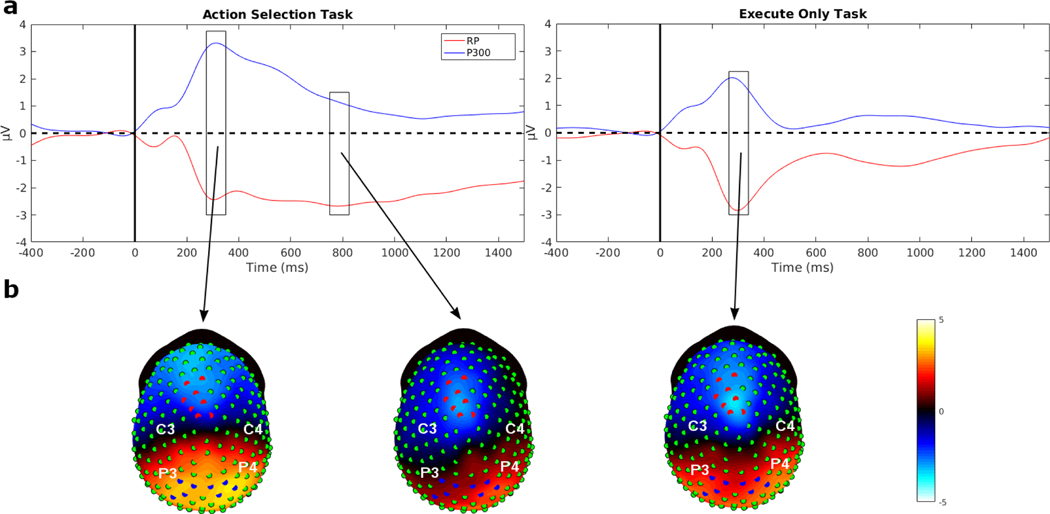
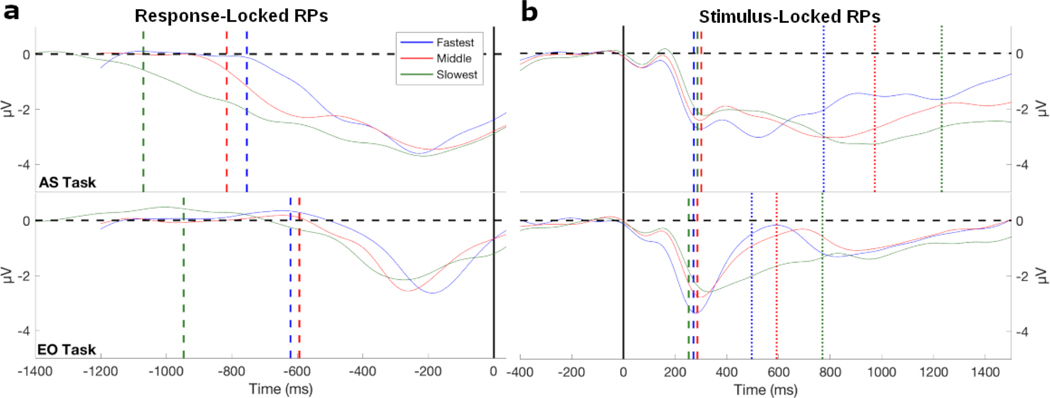
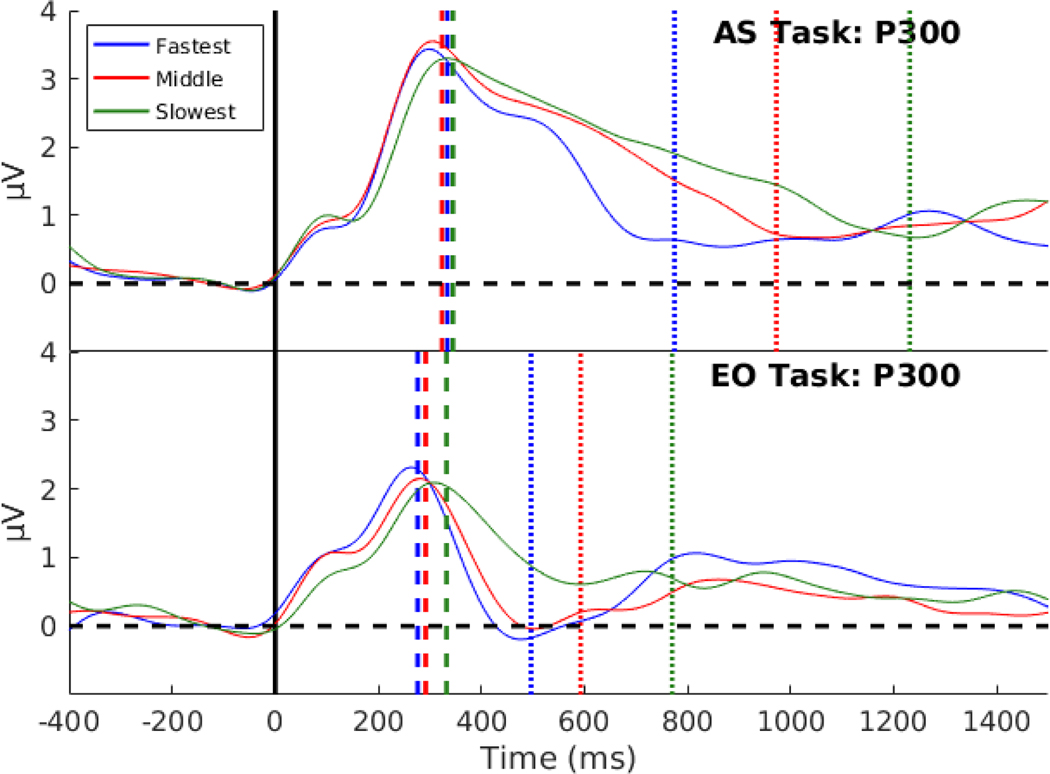
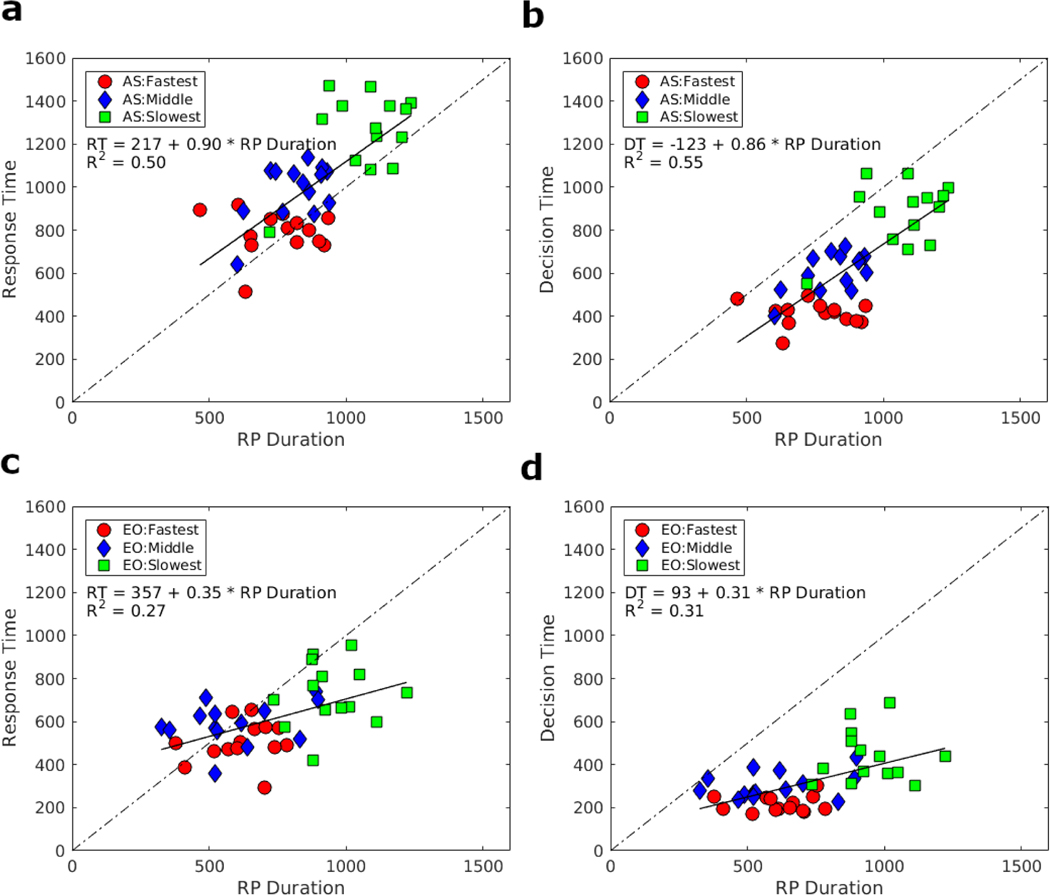
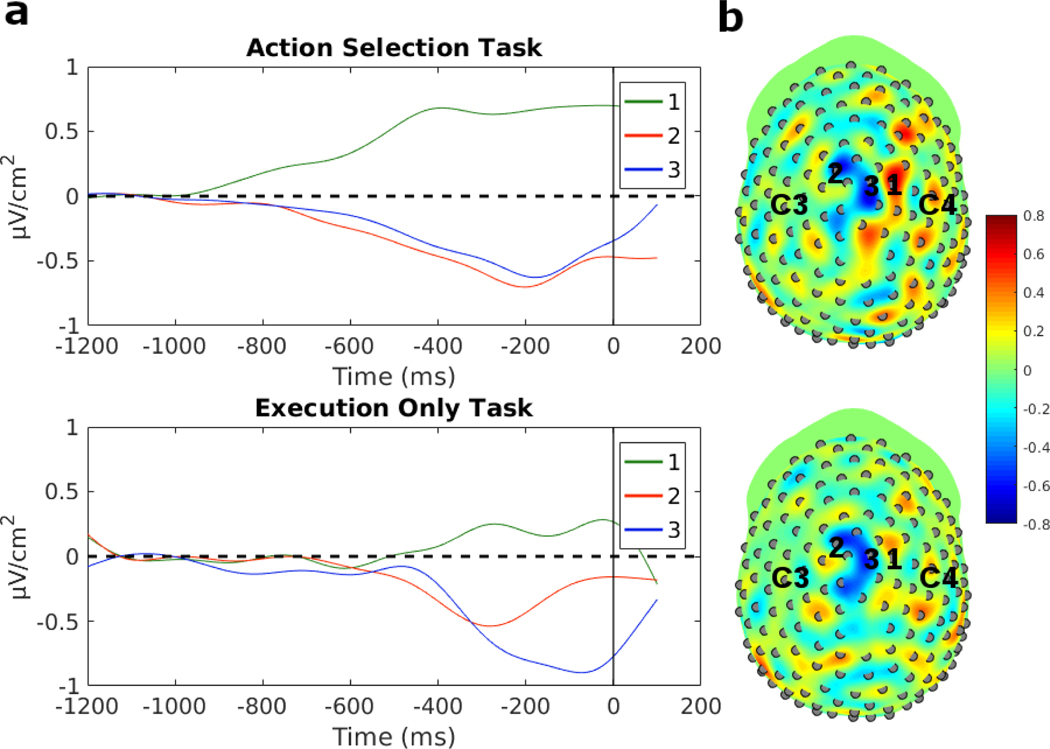
Decision-making in two-alternative forced choice tasks has several underlying components including stimulus encoding, perceptual categorization, response selection, and response execution. Sequential sampling models of decision-making are based on an evidence accumulation process to a decision boundary. Animal and human studies have focused on perceptual categorization and provide evidence linking brain signals in parietal cortex to the evidence accumulation process. In this exploratory study, we use a task where the dominant contribution to response time is response selection and model the response time data with the drift-diffusion model. EEG measurement during the task show that the Readiness Potential (RP) recorded over motor areas has timing consistent with the evidence accumulation process. The duration of the RP predicts decision-making time, the duration of evidence accumulation, suggesting that the RP partly reflects an evidence accumulation process for response selection in the motor system. Thus, evidence accumulation may be a neural implementation of decision-making processes in both perceptual and motor systems. The contributions of perceptual categorization and response selection to evidence accumulation processes in decision-making tasks can be potentially evaluated by examining the timing of perceptual and motor EEG signals.
Keywords: Decision-making; electroencephalography; motor preparation; perceptual categorization; readiness potential; response selection.
Figures