

i-Motif formation and spontaneous deletions in human cells
- PMID: 35253884
- PMCID: PMC8989526
- DOI: 10.1093/nar/gkac158
i-Motif formation and spontaneous deletions in human cells
Abstract
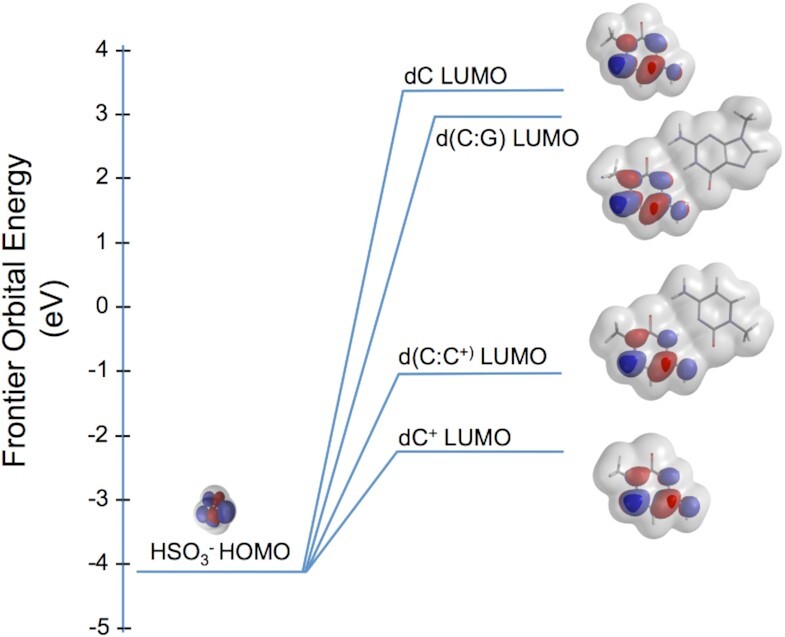
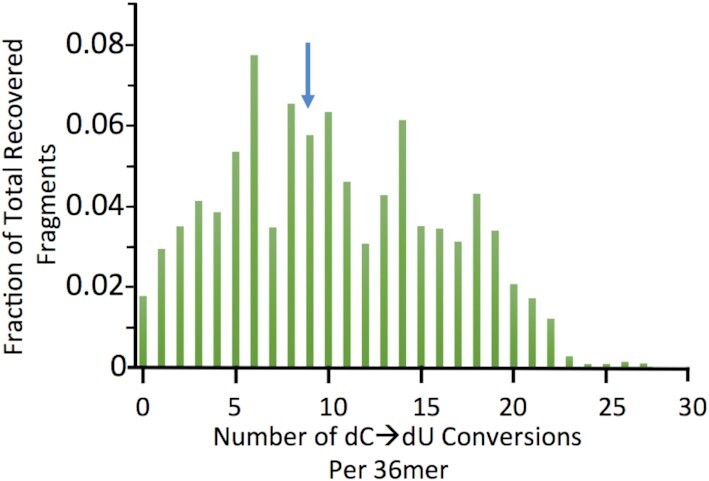
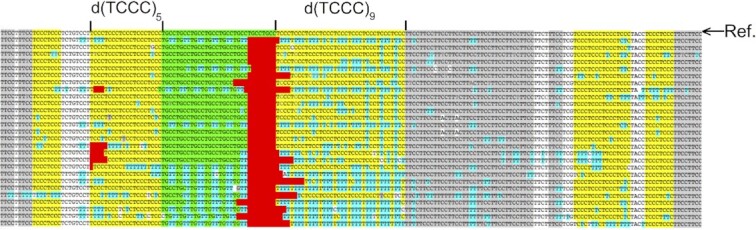
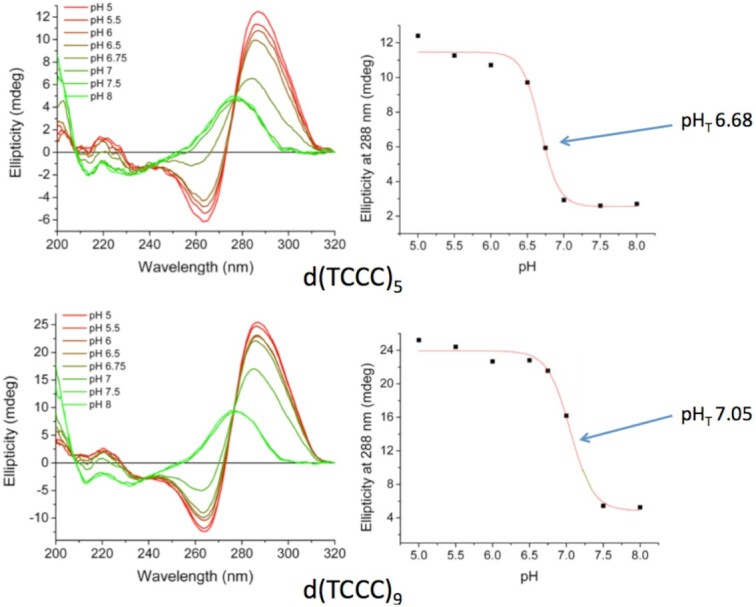
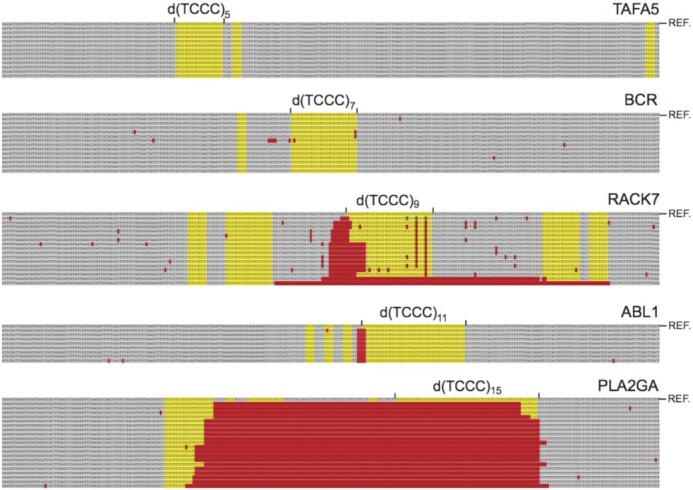
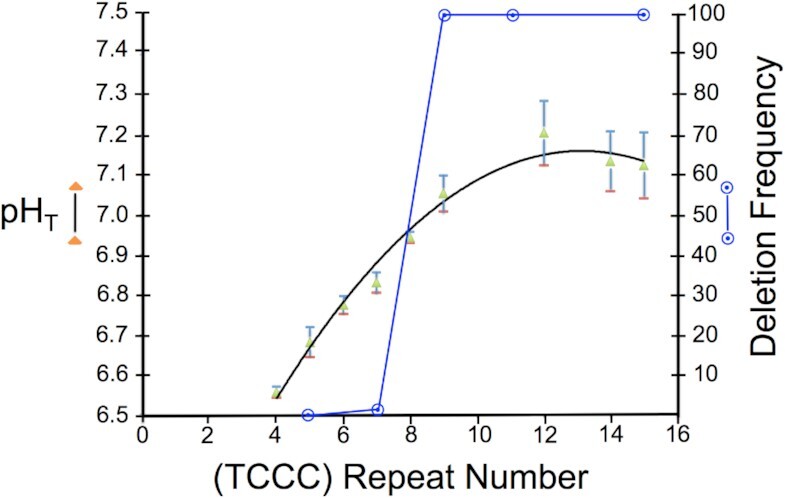
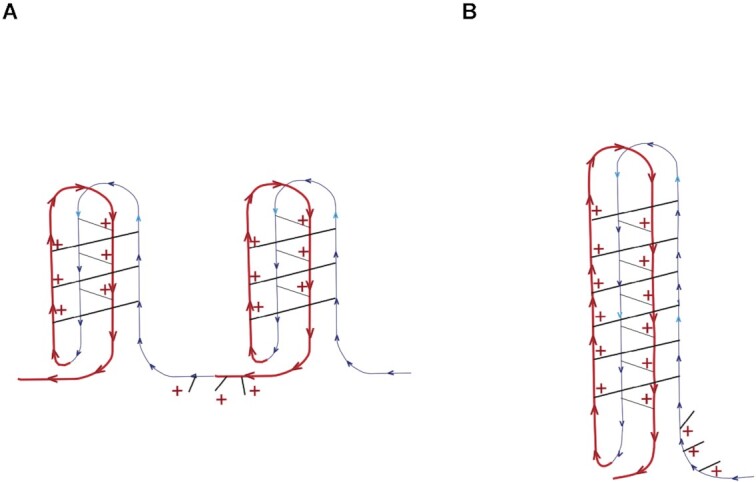
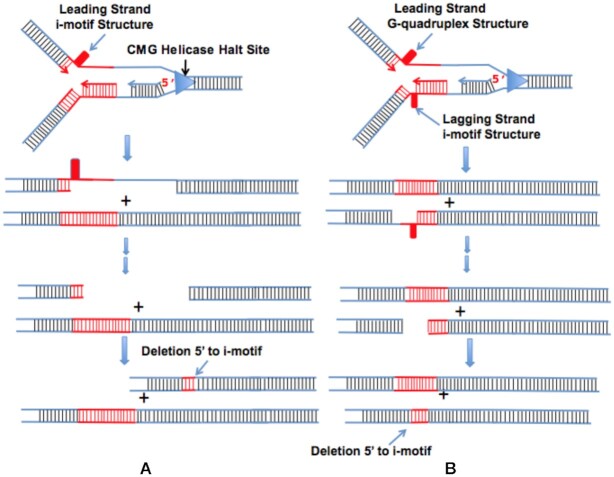
Concatemers of d(TCCC) that were first detected through their association with deletions at the RACK7 locus, are widespread throughout the human genome. Circular dichroism spectra show that d(GGGA)n sequences form G-quadruplexes when n > 3, while i-motif structures form at d(TCCC)n sequences at neutral pH when n ≥ 7 in vitro. In the PC3 cell line, deletions are observed only when the d(TCCC)n variant is long enough to form significant levels of unresolved i-motif structure at neutral pH. The presence of an unresolved i-motif at a representative d(TCCC)n element at RACK7 was suggested by experiments showing that that the region containing the d(TCCC)9 element was susceptible to bisulfite attack in native DNA and that d(TCCC)9 oligo formed an i-motif structure at neutral pH. This in turn suggested that that the i-motif present at this site in native DNA must be susceptible to bisulfite mediated deamination even though it is a closed structure. Bisulfite deamination of the i-motif structure in the model oligodeoxynucleotide was confirmed using mass spectrometry analysis. We conclude that while G-quadruplex formation may contribute to spontaneous mutation at these sites, deletions actually require the potential for i-motif to form and remain unresolved at neutral pH.
© The Author(s) 2022. Published by Oxford University Press on behalf of Nucleic Acids Research.
Figures
References
-
- Sundquist W.I., Klug A.. Telomeric DNA dimerizes by formation of guanine tetrads between hairpin loops. Nature. 1989; 342:825–829. - PubMed
-
- Day H.A., Pavlou P., Waller Z.A.. i-Motif DNA: structure, stability and targeting with ligands. Bioorg. Med. Chem. 2014; 22:4407–4418. - PubMed
-
- Smith S.S. Evolutionary expansion of structurally complex DNA sequences. Cancer Genomics Proteomics. 2010; 7:207–215. - PubMed
