

Functions and consequences of AID/APOBEC-mediated DNA and RNA deamination
- PMID: 35256818
- PMCID: PMC8900473
- DOI: 10.1038/s41576-022-00459-8
Functions and consequences of AID/APOBEC-mediated DNA and RNA deamination
Abstract
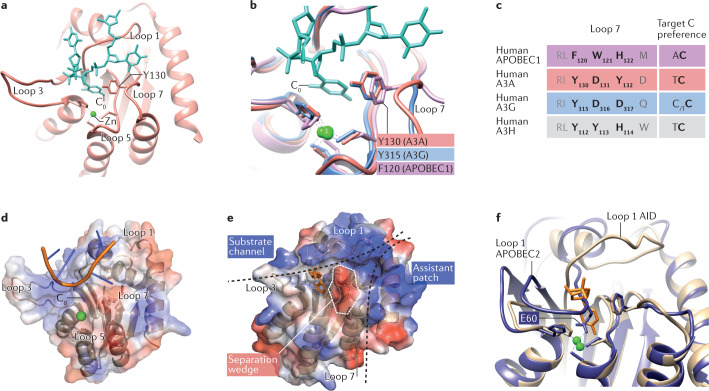
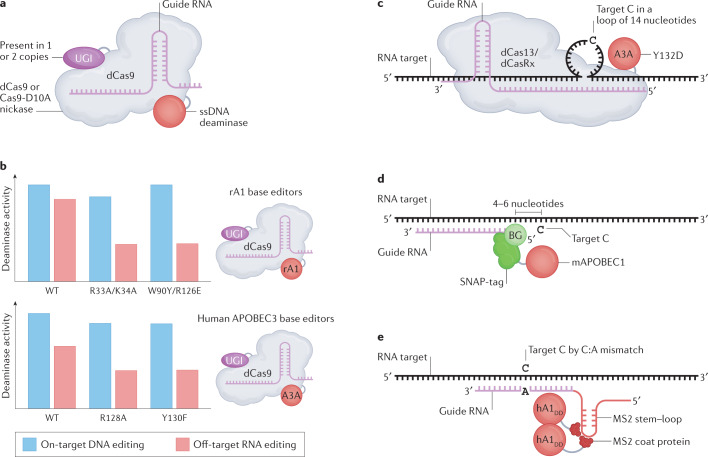
The AID/APOBEC polynucleotide cytidine deaminases have historically been classified as either DNA mutators or RNA editors based on their first identified nucleic acid substrate preference. DNA mutators can generate functional diversity at antibody genes but also cause genomic instability in cancer. RNA editors can generate informational diversity in the transcriptome of innate immune cells, and of cancer cells. Members of both classes can act as antiviral restriction factors. Recent structural work has illuminated differences and similarities between AID/APOBEC enzymes that can catalyse DNA mutation, RNA editing or both, suggesting that the strict functional classification of members of this family should be reconsidered. As many of these enzymes have been employed for targeted genome (or transcriptome) editing, a more holistic understanding will help improve the design of therapeutically relevant programmable base editors.
© 2022. Springer Nature Limited.
Conflict of interest statement
The authors declare no competing interests.
Figures



References
-
- Crick FH. On protein synthesis. Symposia Soc. Exp. Biol. 1958;12:138–163. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
