

Dissecting The role of Plasmodium metacaspase-2 in malaria gametogenesis and sporogony
- PMID: 35264080
- PMCID: PMC8973346
- DOI: 10.1080/22221751.2022.2052357
Dissecting The role of Plasmodium metacaspase-2 in malaria gametogenesis and sporogony
Abstract
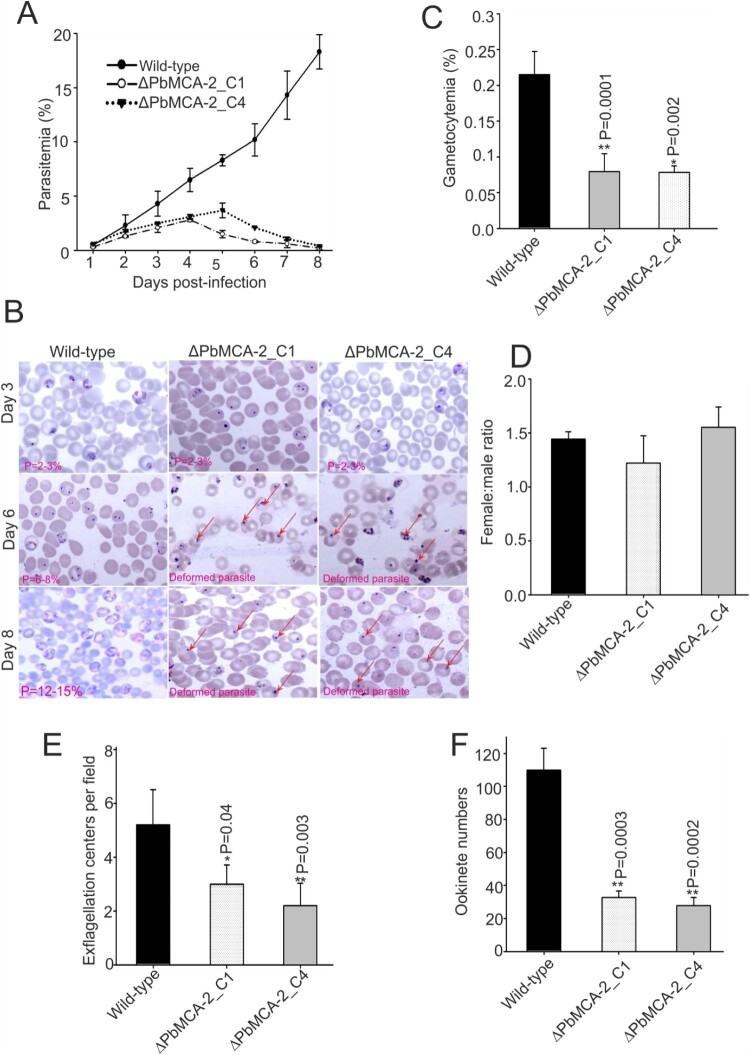
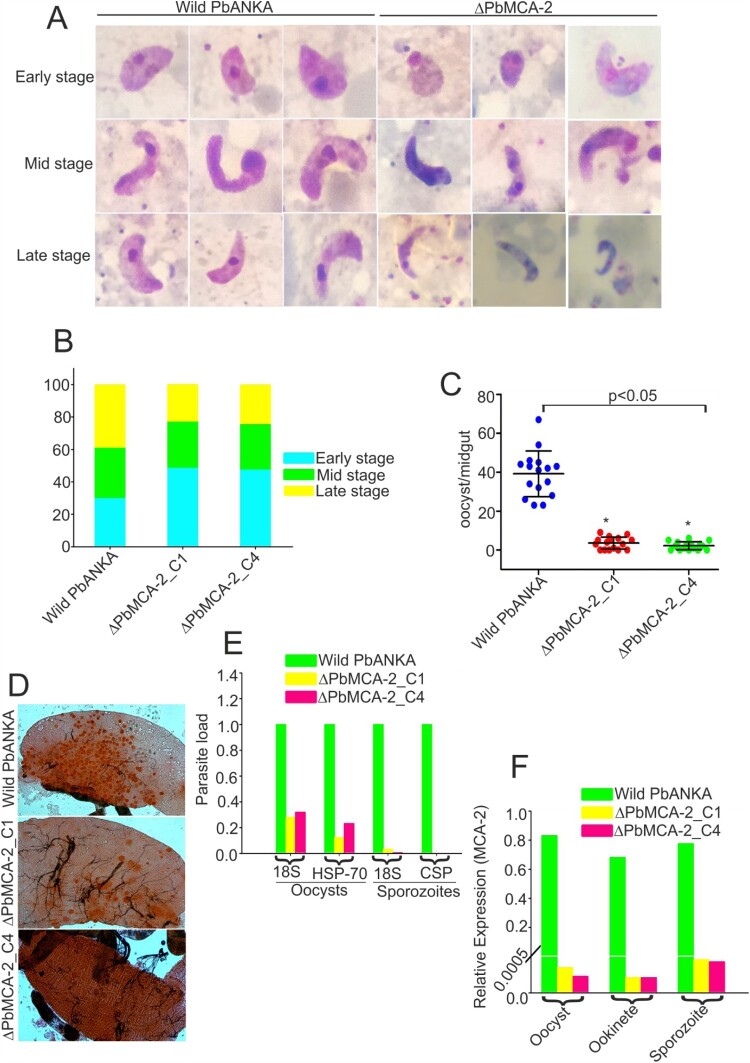
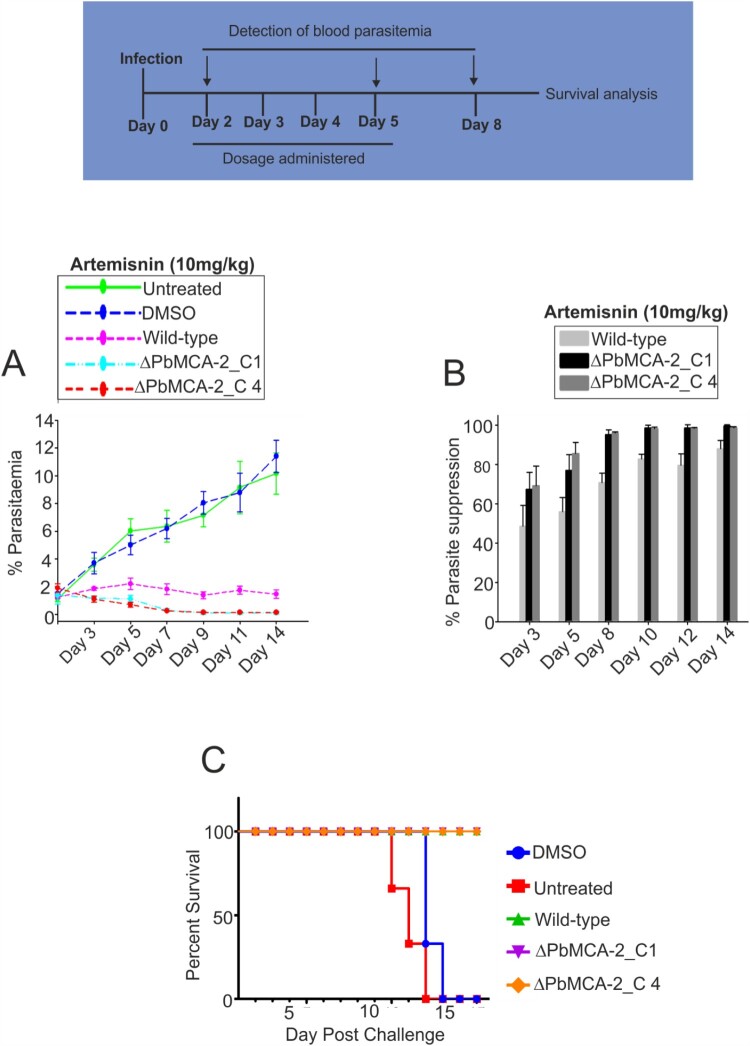
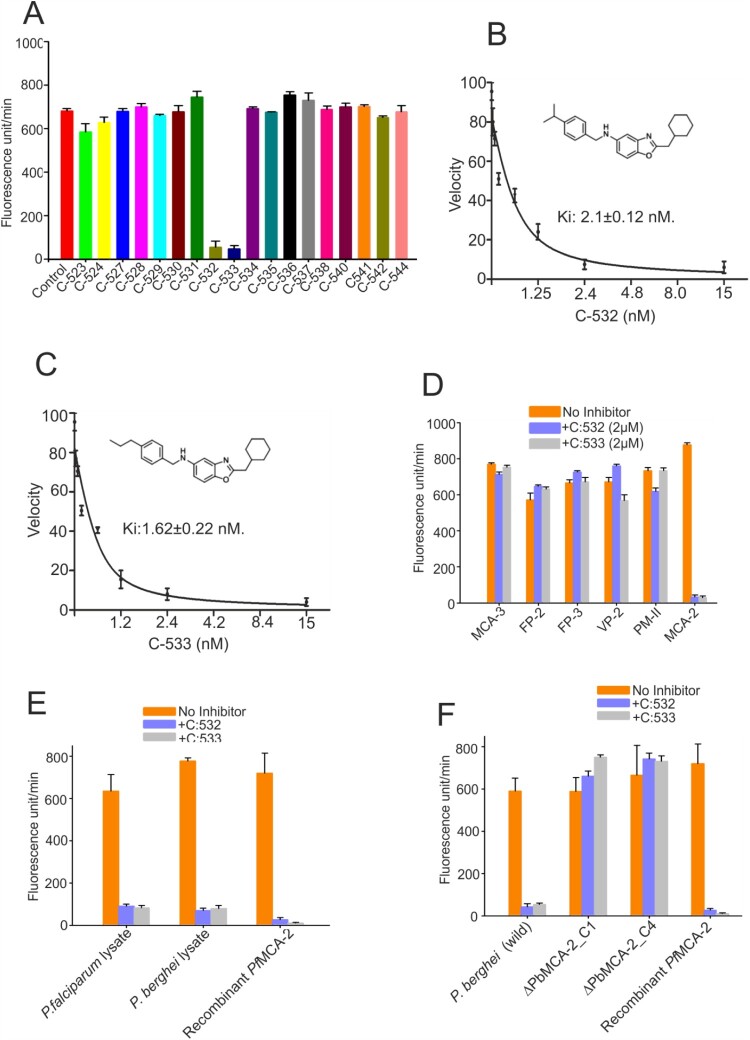
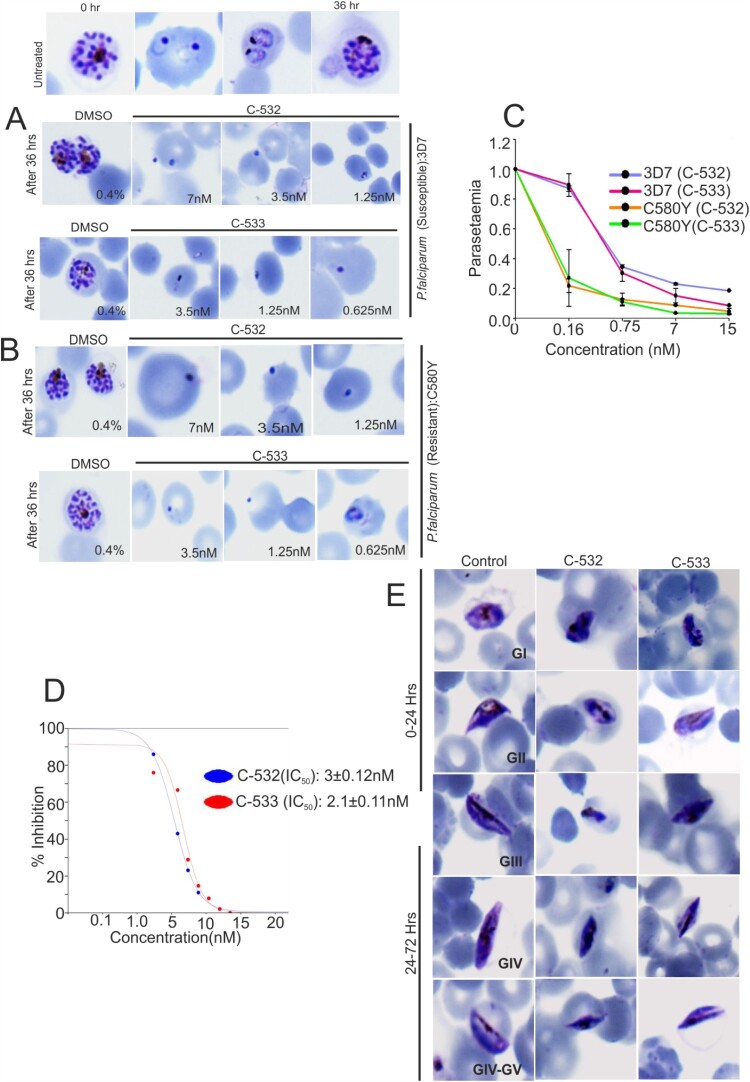
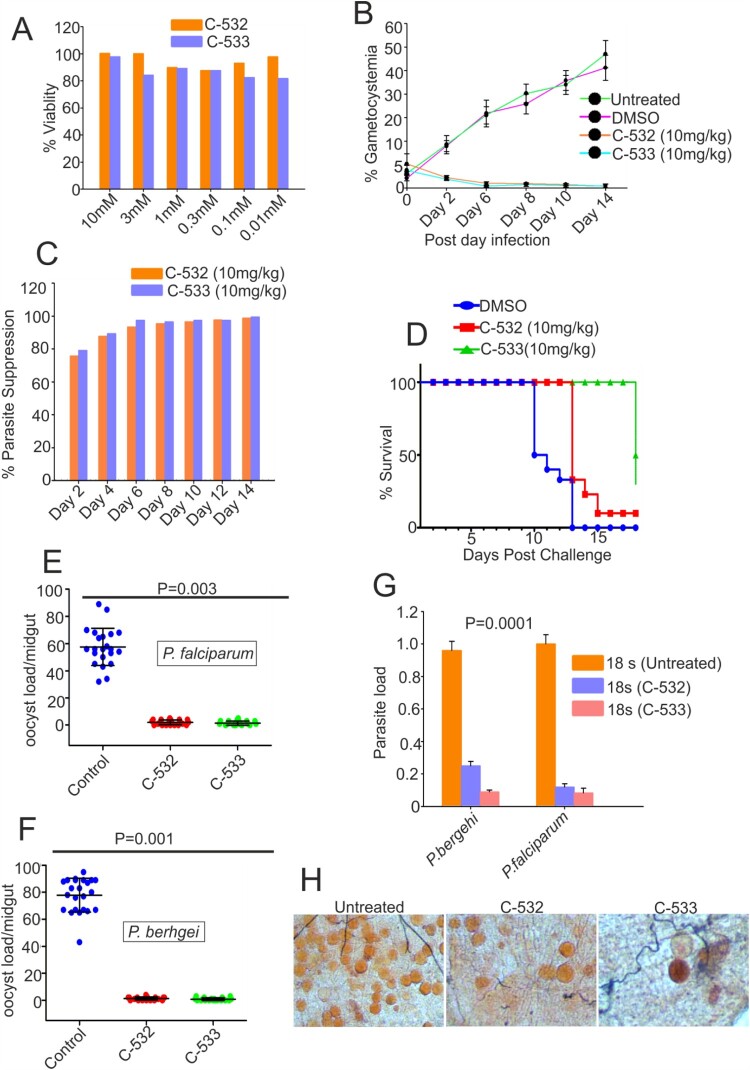
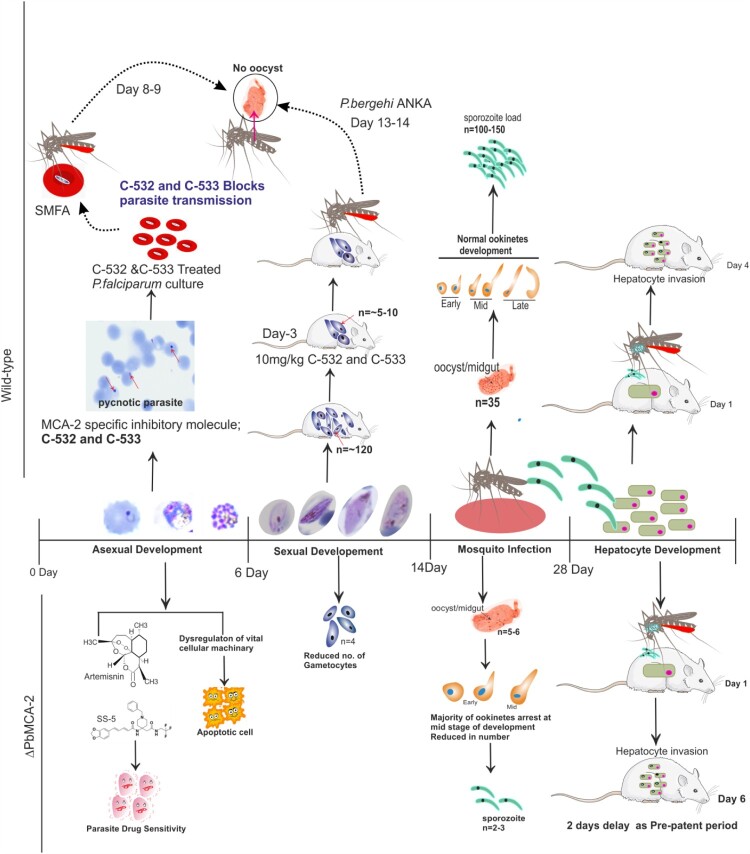
The family of apicomplexan specific proteins contains caspases-like proteins called "metacaspases". These enzymes are present in the malaria parasite but absent in human; therefore, these can be explored as potential drug targets. We deleted the MCA-2 gene from Plasmodium berghei genome using a gene knockout strategy to decipher its precise function. This study has identified that MCA-2 plays an important role in parasite transmission since it is critical for the formation of gametocytes and for maintaining an appropriate number of infectious sporozoites required for sporogony. It is noticeable that a significant reduction in gametocyte, oocysts, ookinete and sporozoites load along with a delay in hepatocytes invasion were observed in the MCA-2 knockout parasite. Furthermore, a study found the two MCA-2 inhibitory molecules known as C-532 and C-533, which remarkably inhibited the MCA-2 activity, abolished the in vitro parasite growth, and also impaired the transmission cycle of P. falciparum and P. berghei in An. stephensi. Our findings indicate that the deletion of MCA-2 hampers the Plasmodium development during erythrocytic and exo-erythrocytic stages, and its inhibition by C-532 and C-533 critically affects the malaria transmission biology.
Keywords: An. stephensi; Malaria; Plasmodium transmission; gametogenesis; metacaspase; ookinete; specific inhibitor; sporogony.
Conflict of interest statement
No potential conflict of interest was reported by the author(s).
Figures
Similar articles
-
A G-Protein-Coupled Receptor Modulates Gametogenesis via PKG-Mediated Signaling Cascade in Plasmodium berghei.Microbiol Spectr. 2022 Apr 27;10(2):e0015022. doi: 10.1128/spectrum.00150-22. Epub 2022 Apr 11. Microbiol Spectr. 2022. PMID: 35404079 Free PMC article.
-
A conserved malaria parasite antigen Pb22 plays a critical role in male gametogenesis in Plasmodium berghei.Cell Microbiol. 2021 Mar;23(3):e13294. doi: 10.1111/cmi.13294. Epub 2020 Dec 10. Cell Microbiol. 2021. PMID: 33222390 Free PMC article.
-
Dysregulated gene expression in oocysts of Plasmodium berghei LAP mutants.Mol Biochem Parasitol. 2019 Apr;229:1-5. doi: 10.1016/j.molbiopara.2019.02.001. Epub 2019 Feb 10. Mol Biochem Parasitol. 2019. PMID: 30753856 Free PMC article.
-
Plasmodium falciparum gametocytes: still many secrets of a hidden life.Mol Microbiol. 2007 Oct;66(2):291-302. doi: 10.1111/j.1365-2958.2007.05904.x. Epub 2007 Sep 3. Mol Microbiol. 2007. PMID: 17784927 Review.
-
Gametogenesis in Plasmodium: Delving Deeper to Connect the Dots.Front Cell Infect Microbiol. 2022 Jun 15;12:877907. doi: 10.3389/fcimb.2022.877907. eCollection 2022. Front Cell Infect Microbiol. 2022. PMID: 35782151 Free PMC article. Review.
Cited by
-
The Kelch13 compartment contains highly divergent vesicle trafficking proteins in malaria parasites.PLoS Pathog. 2023 Dec 1;19(12):e1011814. doi: 10.1371/journal.ppat.1011814. eCollection 2023 Dec. PLoS Pathog. 2023. PMID: 38039338 Free PMC article.
-
Cell Death of P. vivax Blood Stages Occurs in Absence of Classical Apoptotic Events and Induces Eryptosis of Parasitized Host Cells.Pathogens. 2024 Aug 9;13(8):673. doi: 10.3390/pathogens13080673. Pathogens. 2024. PMID: 39204273 Free PMC article.
-
Drug-induced ER stress leads to induction of programmed cell death pathways of the malaria parasite.Parasitol Res. 2024 Jul 8;123(7):263. doi: 10.1007/s00436-024-08281-3. Parasitol Res. 2024. PMID: 38976068 Free PMC article.
-
Genomics of Plasmodium vivax in Colombia reveals evidence of local bottle-necking and inter-country connectivity in the Americas.Sci Rep. 2023 Nov 13;13(1):19779. doi: 10.1038/s41598-023-46076-1. Sci Rep. 2023. PMID: 37957271 Free PMC article.
-
Structure-function study of a Ca2+-independent metacaspase involved in lateral root emergence.Proc Natl Acad Sci U S A. 2023 May 30;120(22):e2303480120. doi: 10.1073/pnas.2303480120. Epub 2023 May 22. Proc Natl Acad Sci U S A. 2023. PMID: 37216519 Free PMC article.
References
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical