

Mice recognize 3D objects from recalled 2D pictures, support for picture-object equivalence
- PMID: 35264621
- PMCID: PMC8907285
- DOI: 10.1038/s41598-022-07782-4
Mice recognize 3D objects from recalled 2D pictures, support for picture-object equivalence
Abstract
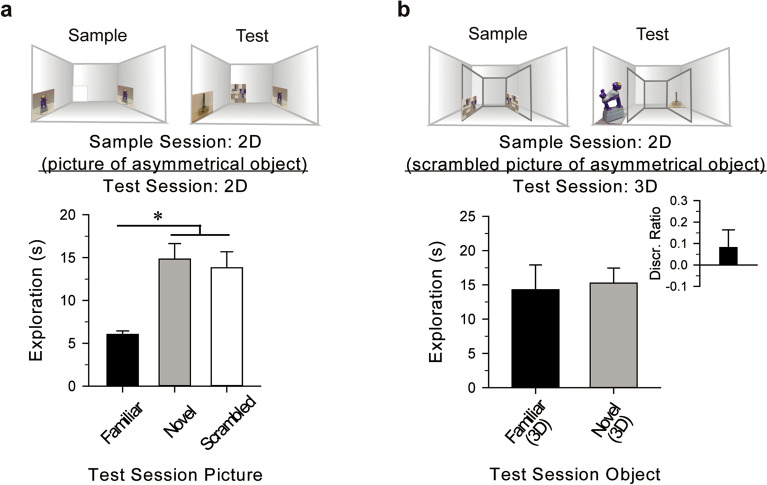
Picture-object equivalence or recognizing a three-dimensional (3D) object after viewing a two-dimensional (2D) photograph of that object, is a higher-order form of visual cognition that may be beyond the perceptual ability of rodents. Behavioral and neurobiological mechanisms supporting picture-object equivalence are not well understood. We used a modified visual recognition memory task, reminiscent of those used for primates, to test whether picture-object equivalence extends to mice. Mice explored photographs of an object during a sample session, and 24 h later were presented with the actual 3D object from the photograph and a novel 3D object, or the stimuli were once again presented in 2D form. Mice preferentially explored the novel stimulus, indicating recognition of the "familiar" stimulus, regardless of whether the sample photographs depicted radially symmetric or asymmetric, similar, rotated, or abstract objects. Discrimination did not appear to be guided by individual object features or low-level visual stimuli. Inhibition of CA1 neuronal activity in dorsal hippocampus impaired discrimination, reflecting impaired memory of the 2D sample object. Collectively, results from a series of experiments provide strong evidence that picture-object equivalence extends to mice and is hippocampus-dependent, offering important support for the appropriateness of mice for investigating mechanisms of human cognition.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures





References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
