

Central amygdala corticotropin-releasing factor neurons promote hyponeophagia but do not control alcohol drinking in mice
- PMID: 35264727
- PMCID: PMC9149056
- DOI: 10.1038/s41380-022-01496-9
Central amygdala corticotropin-releasing factor neurons promote hyponeophagia but do not control alcohol drinking in mice
Abstract
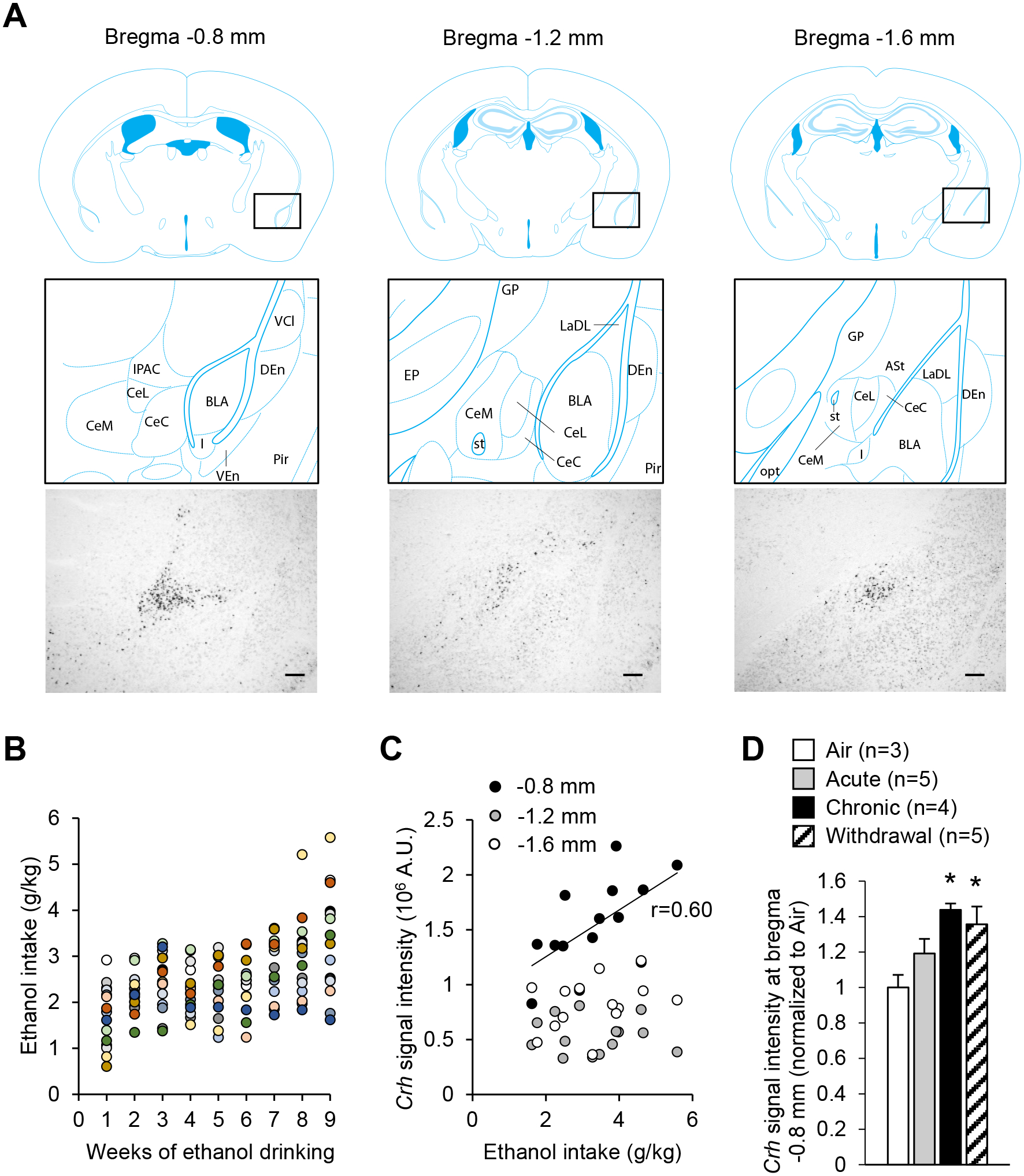
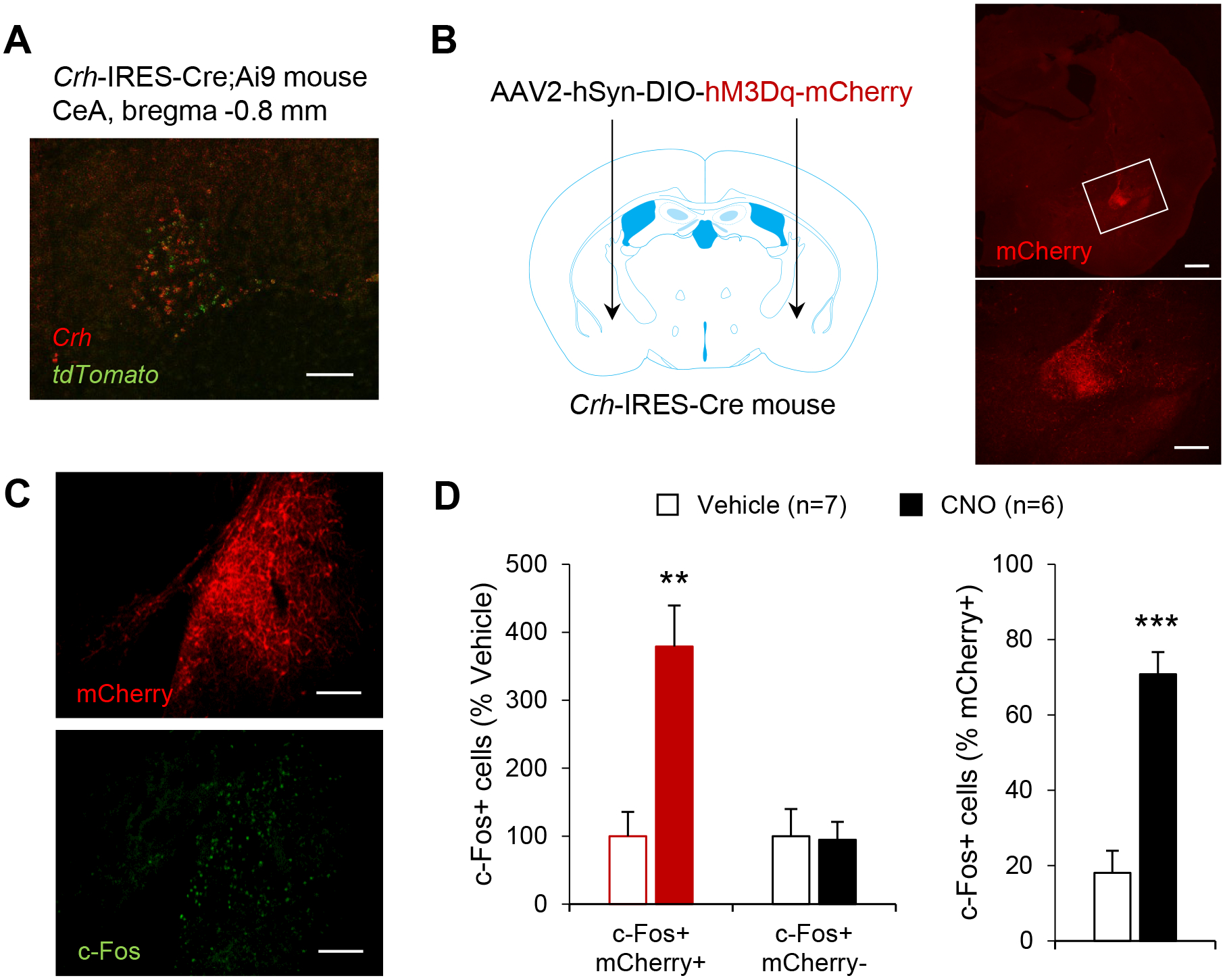
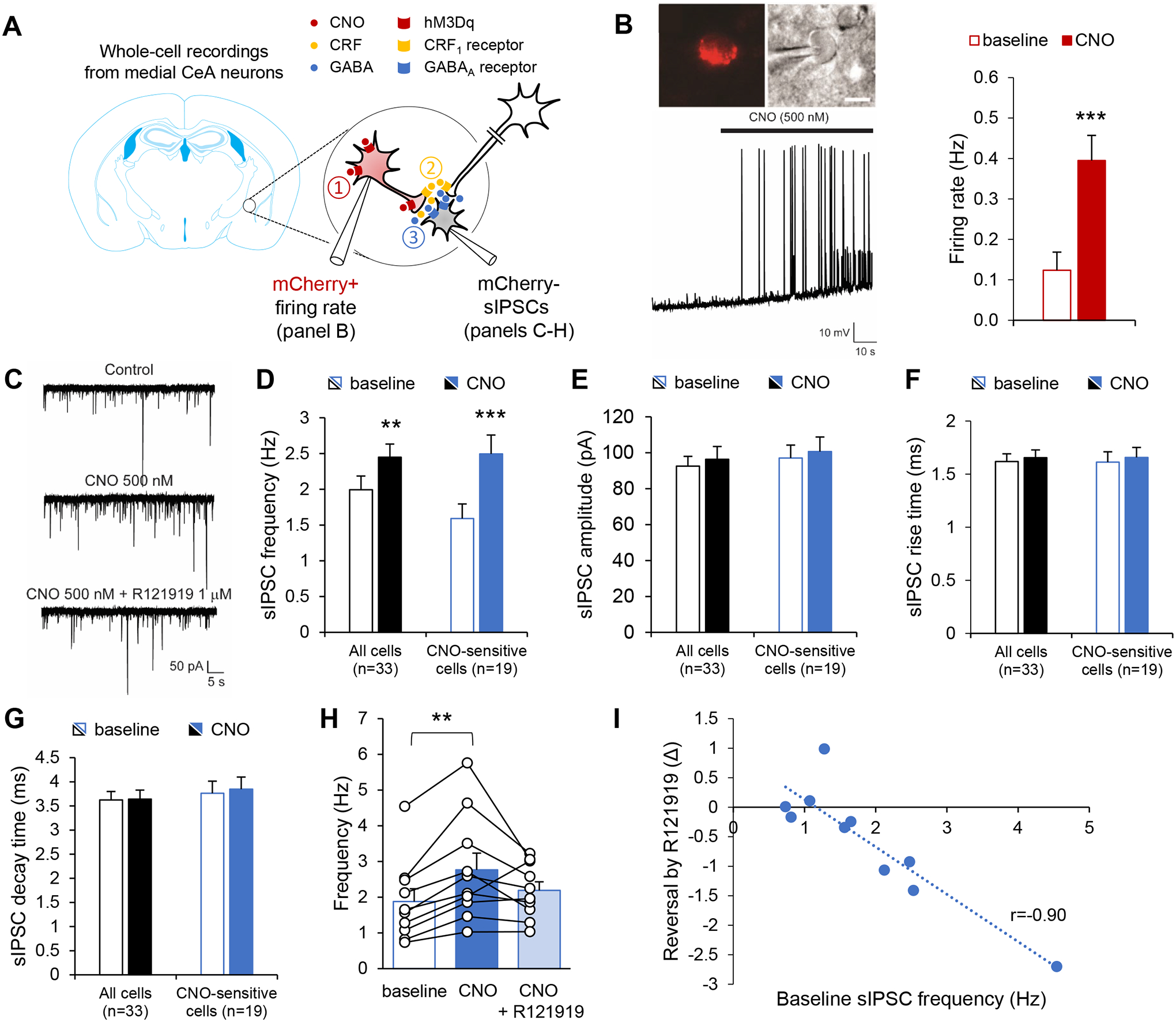
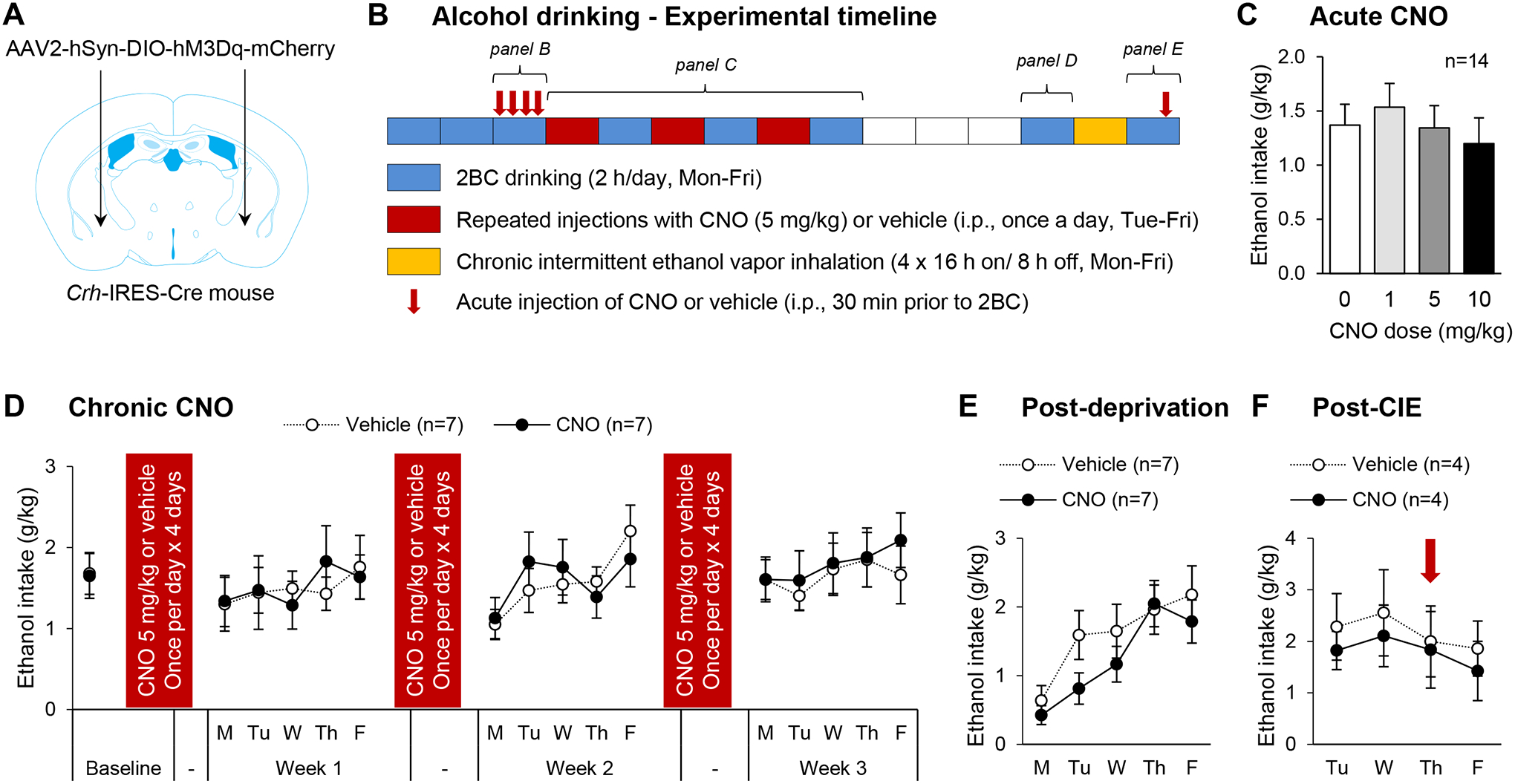
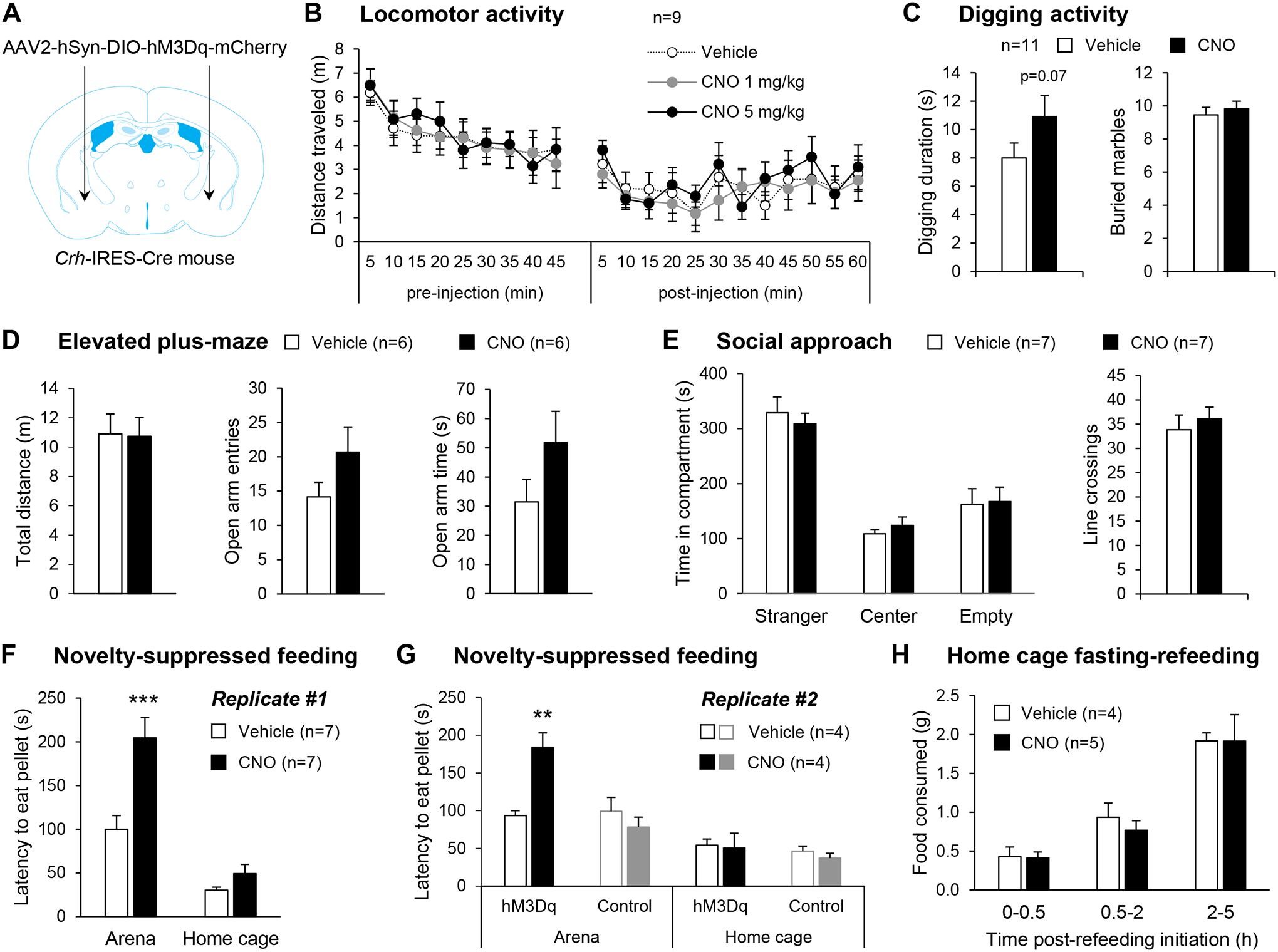
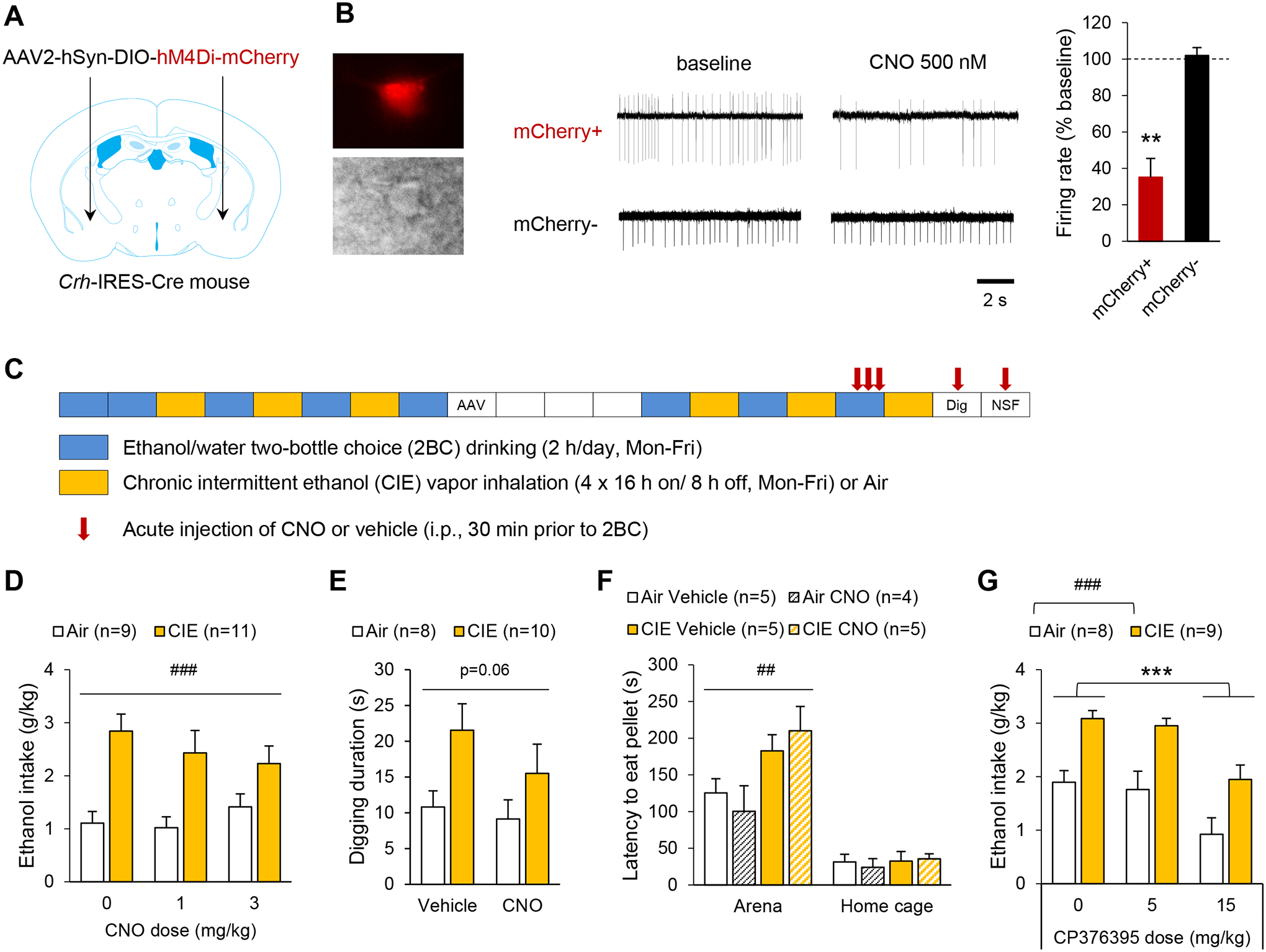
Corticotropin-releasing factor (CRF) signaling in the central nucleus of the amygdala (CeA) plays a critical role in rodent models of excessive alcohol drinking. However, the source of CRF acting in the CeA during alcohol withdrawal remains to be identified. In the present study, we hypothesized that CeA CRF interneurons may represent a behaviorally relevant source of CRF to the CeA increasing motivation for alcohol via negative reinforcement. We first observed that Crh mRNA expression in the anterior part of the mouse CeA correlates positively with alcohol intake in C57BL/6J males with a history of chronic binge drinking followed by abstinence and increases upon exposure to chronic intermittent ethanol (CIE) vapor inhalation. We then found that chemogenetic activation of CeA CRF neurons in Crh-IRES-Cre mouse brain slices increases gamma-aminobutyric acid (GABA) release in the medial CeA, in part via CRF1 receptor activation. While chemogenetic stimulation exacerbated novelty-induced feeding suppression (NSF) in alcohol-naïve mice, thereby mimicking the effect of withdrawal from CIE, it had no effect on voluntary alcohol consumption, following either acute or chronic manipulation. Furthermore, chemogenetic inhibition of CeA CRF neurons did not affect alcohol consumption or NSF in chronic alcohol drinkers exposed to air or CIE. Altogether, these findings indicate that CeA CRF neurons produce local release of GABA and CRF and promote hyponeophagia in naïve mice, but do not drive alcohol intake escalation or negative affect in CIE-withdrawn mice. The latter result contrasts with previous findings in rats and demonstrates species specificity of CRF circuit engagement in alcohol dependence.
© 2022. The Author(s), under exclusive licence to Springer Nature Limited.
Conflict of interest statement
Disclosure
The authors have no conflicts of interest to declare.
Figures
References
-
- National Survey on Drug Use and Health: Substance Abuse and Mental Health Services Administration; 2015. - PubMed
-
- Koob GF, Le Moal M. Addiction and the brain antireward system. Annual review of psychology 2008; 59: 29–53. - PubMed
-
- Becker HC, Lopez MF. Increased ethanol drinking after repeated chronic ethanol exposure and withdrawal experience in C57BL/6 mice. Alcohol Clin Exp Res 2004; 28(12): 1829–1838. - PubMed
-
- Rhodes JS, Best K, Belknap JK, Finn DA, Crabbe JC. Evaluation of a simple model of ethanol drinking to intoxication in C57BL/6J mice. Physiology & behavior 2005; 84(1): 53–63. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- U01 AA013498/AA/NIAAA NIH HHS/United States
- R33 AA027636/AA/NIAAA NIH HHS/United States
- R21 AA024198/AA/NIAAA NIH HHS/United States
- R21 AA027372/AA/NIAAA NIH HHS/United States
- K99 AA023002/AA/NIAAA NIH HHS/United States
- P60 AA006420/AA/NIAAA NIH HHS/United States
- R01 AA015566/AA/NIAAA NIH HHS/United States
- F32 AA024952/AA/NIAAA NIH HHS/United States
- P50 AA006420/AA/NIAAA NIH HHS/United States
- R00 AA023002/AA/NIAAA NIH HHS/United States
- R01 AA026685/AA/NIAAA NIH HHS/United States
- R21 AA027636/AA/NIAAA NIH HHS/United States
- R01 AA021491/AA/NIAAA NIH HHS/United States
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
