

The Amino Terminal Domain and Modulation of Connexin36 Gap Junction Channels by Intracellular Magnesium Ions
- PMID: 35264979
- PMCID: PMC8899287
- DOI: 10.3389/fphys.2022.839223
The Amino Terminal Domain and Modulation of Connexin36 Gap Junction Channels by Intracellular Magnesium Ions
Abstract
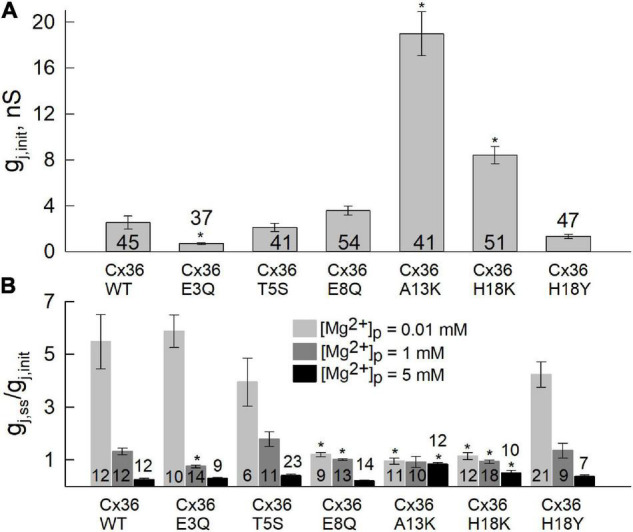
Electrical synapses between neurons in the mammalian CNS are predominantly formed of the connexin36 (Cx36) gap junction (GJ) channel protein. Unique among GJs formed of a number of other members of the Cx gene family, Cx36 GJs possess a high sensitivity to intracellular Mg2+ that can robustly act to modulate the strength of electrical synaptic transmission. Although a putative Mg2+ binding site was previously identified to reside in the aqueous pore in the first extracellular (E1) loop domain, the involvement of the N-terminal (NT) domain in the atypical response of Cx36 GJs to pH was shown to depend on intracellular levels of Mg2+. In this study, we examined the impact of amino acid substitutions in the NT domain on Mg2+ modulation of Cx36 GJs, focusing on positions predicted to line the pore funnel, which constitutes the cytoplasmic entrance of the channel pore. We find that charge substitutions at the 8th, 13th, and 18th positions had pronounced effects on Mg2+ sensitivity, particularly at position 13 at which an A13K substitution completely abolished sensitivity to Mg2+. To assess potential mechanisms of Mg2+ action, we constructed and tested a series of mathematical models that took into account gating of the component hemichannels in a Cx36 GJ channel as well as Mg2+ binding to each hemichannel in open and/or closed states. Simultaneous model fitting of measurements of junctional conductance, gj, and transjunctional Mg2+ fluxes using a fluorescent Mg2+ indicator suggested that the most viable mechanism for Cx36 regulation by Mg2+ entails the binding of Mg2+ to and subsequent stabilization of the closed state in each hemichannel. Reduced permeability to Mg2+ was also evident, particularly for the A13K substitution, but homology modeling of all charge-substituted NT variants showed only a moderate correlation between a reduction in the negative electrostatic potential and a reduction in the permeability to Mg2+ ions. Given the reported role of the E1 domain in Mg2+ binding together with the impact of NT substitutions on gating and the apparent state-dependence of Mg2+ binding, this study suggests that the NT domain can be an integral part of Mg2+ modulation of Cx36 GJs likely through the coupling of conformational changes between NT and E1 domains.
Keywords: connexin; electrophysiology; gap junction; intracellular Mg2+; ion permeability.
Copyright © 2022 Kraujalis, Gudaitis, Kraujaliene, Snipas, Palacios-Prado and Verselis.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures





Similar articles
-
The N-terminal domain and voltage dependence of connexin-36 gap junction channels.Arch Biochem Biophys. 2025 Oct;772:110556. doi: 10.1016/j.abb.2025.110556. Epub 2025 Jul 19. Arch Biochem Biophys. 2025. PMID: 40692001
-
Emerging issues of connexin channels: biophysics fills the gap.Q Rev Biophys. 2001 Aug;34(3):325-472. doi: 10.1017/s0033583501003705. Q Rev Biophys. 2001. PMID: 11838236 Review.
-
Intracellular magnesium-dependent modulation of gap junction channels formed by neuronal connexin36.J Neurosci. 2013 Mar 13;33(11):4741-53. doi: 10.1523/JNEUROSCI.2825-12.2013. J Neurosci. 2013. PMID: 23486946 Free PMC article.
-
Variants with increased negative electrostatic potential in the Cx50 gap junction pore increased unitary channel conductance and magnesium modulation.Biochem J. 2018 Nov 6;475(21):3315-3330. doi: 10.1042/BCJ20180523. Biochem J. 2018. PMID: 30287491
-
The contribution of electrical synapses to field potential oscillations in the hippocampal formation.Front Neural Circuits. 2014 Apr 3;8:32. doi: 10.3389/fncir.2014.00032. eCollection 2014. Front Neural Circuits. 2014. PMID: 24772068 Free PMC article. Review.
Cited by
-
Connexin Gap Junction Channels and Hemichannels: Insights from High-Resolution Structures.Biology (Basel). 2024 Apr 26;13(5):298. doi: 10.3390/biology13050298. Biology (Basel). 2024. PMID: 38785780 Free PMC article. Review.
-
Whole exome sequencing identified mutations of forkhead box I 1 (FOXI1), keratin 6 C (KRT6C) and gap junction protein delta 2 (GJD2) in a low-grade oncocytic tumor of the kidney: a case report.Diagn Pathol. 2025 Feb 21;20(1):21. doi: 10.1186/s13000-025-01616-3. Diagn Pathol. 2025. PMID: 39980061 Free PMC article.
-
A pore locus in the E1 domain differentially regulates Cx26 and Cx30 hemichannel function.J Gen Physiol. 2024 Nov 4;156(11):e202313502. doi: 10.1085/jgp.202313502. Epub 2024 Sep 20. J Gen Physiol. 2024. PMID: 39302316 Free PMC article.
-
Cryo-EM structures of human Cx36/GJD2 neuronal gap junction channel.Nat Commun. 2023 Mar 11;14(1):1347. doi: 10.1038/s41467-023-37040-8. Nat Commun. 2023. PMID: 36906653 Free PMC article.
-
Modeling and analysis of voltage gating of gap junction channels at a single-channel level.Biophys J. 2023 Nov 7;122(21):4176-4193. doi: 10.1016/j.bpj.2023.09.015. Epub 2023 Sep 27. Biophys J. 2023. PMID: 37766427 Free PMC article.
References
-
- Ainscow E. K., Mirshamsi S., Tang T., Ashford M. L., Rutter G. A. (2002). Dynamic imaging of free cytosolic ATP concentration during fuel sensing by rat hypothalamic neurones: evidence for ATP-independent control of ATP-sensitive K(+) channels. J. Physiol. 544 429–445. 10.1113/jphysiol.2002.022434 - DOI - PMC - PubMed
-
- Andrasi E., Igaz S., Molnar Z., Mako S. (2000). Disturbances of magnesium concentrations in various brain areas in Alzheimer’s disease. Magnes. Res. 13 189–196. - PubMed
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous