

Distinct Roles of Honeybee Gut Bacteria on Host Metabolism and Neurological Processes
- PMID: 35266810
- PMCID: PMC9045319
- DOI: 10.1128/spectrum.02438-21
Distinct Roles of Honeybee Gut Bacteria on Host Metabolism and Neurological Processes
Abstract
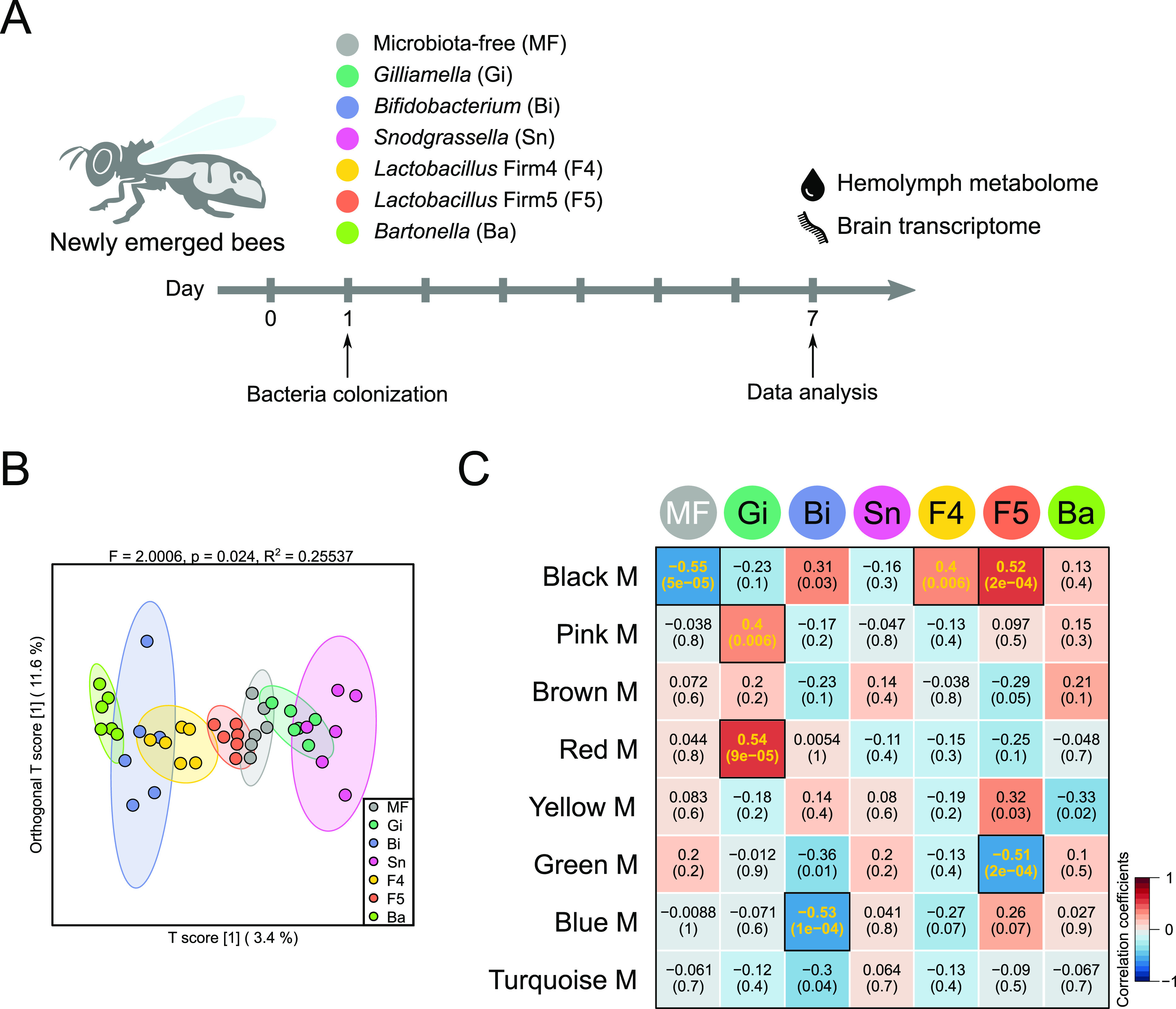
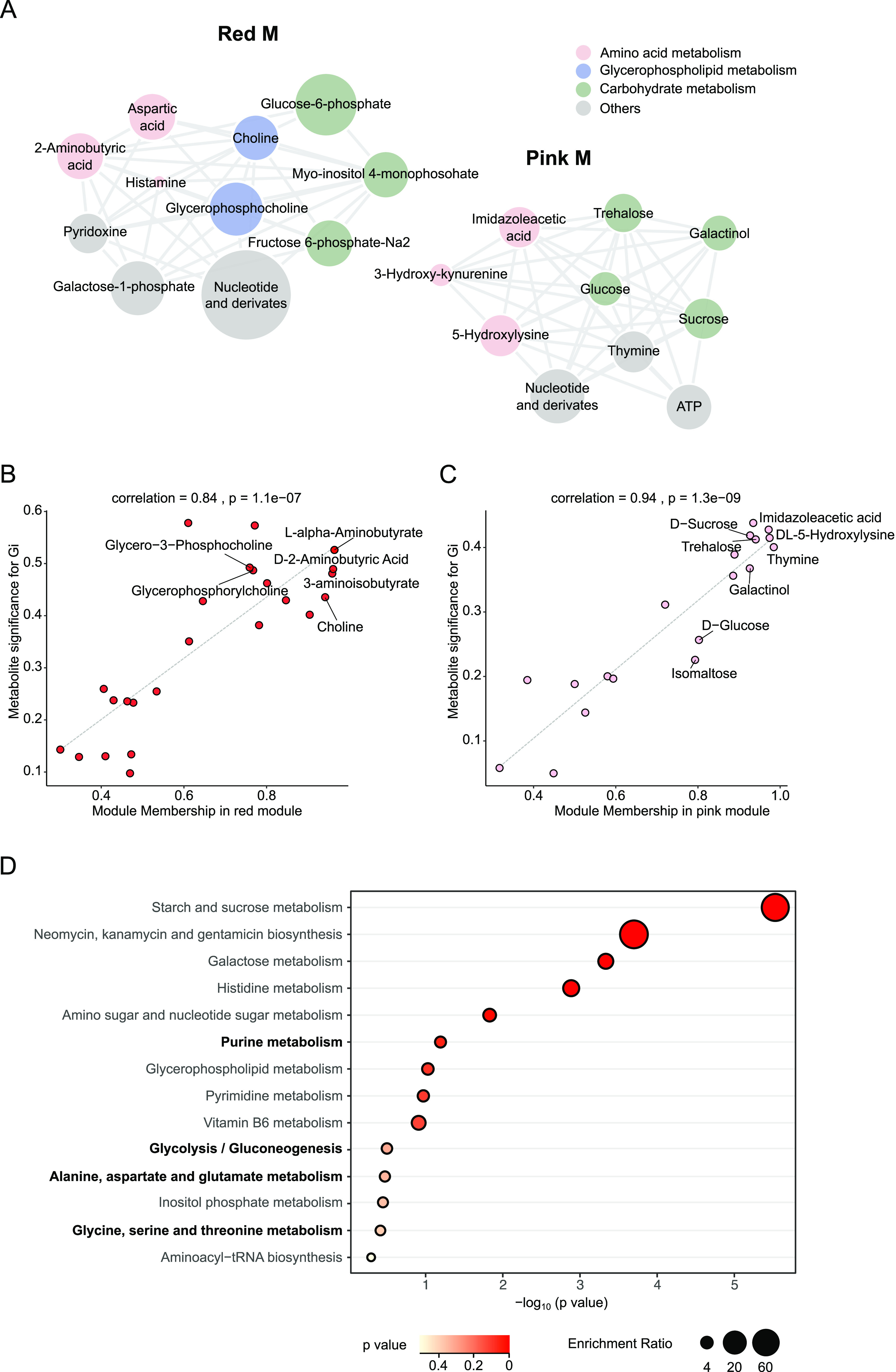
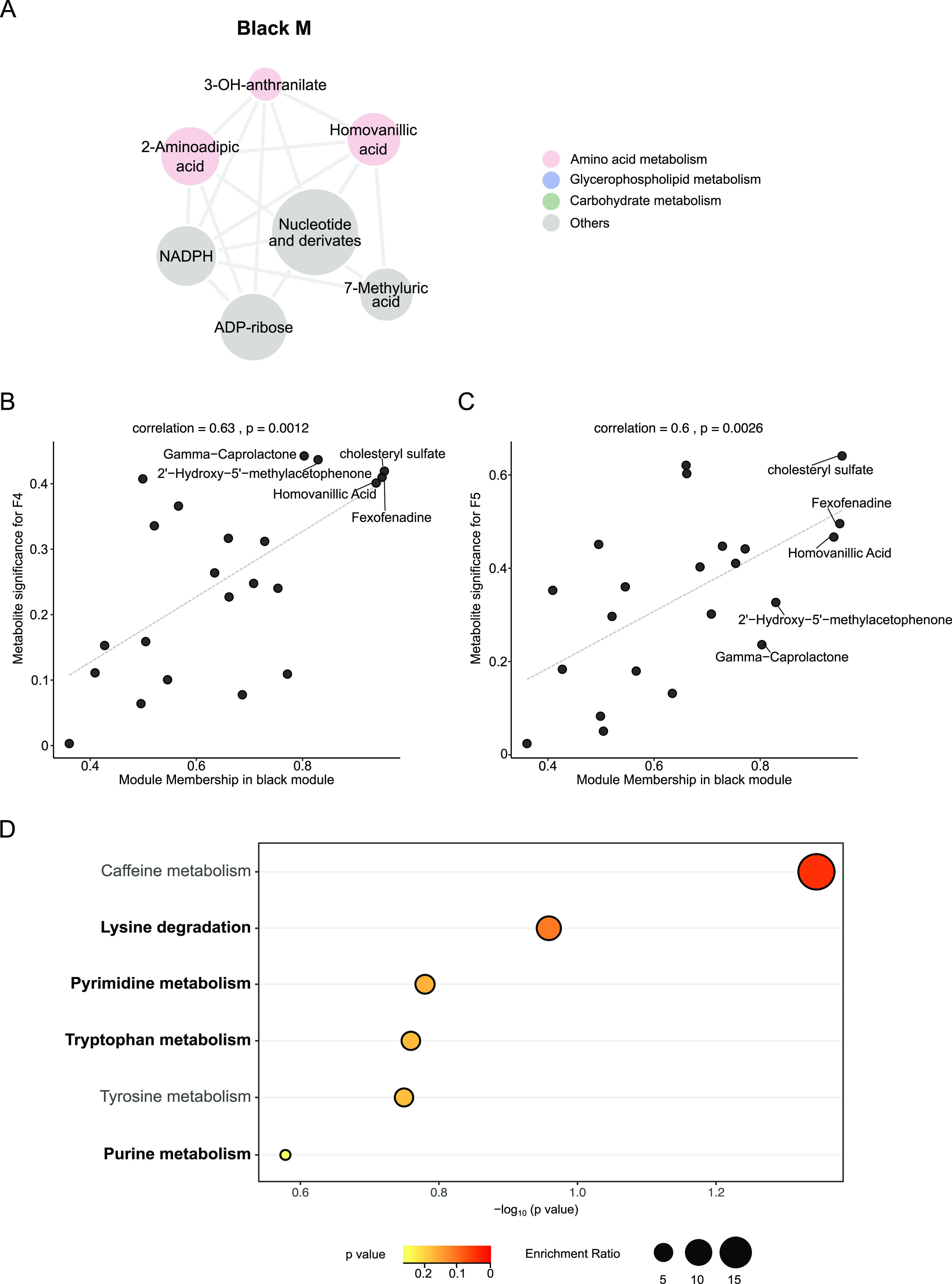
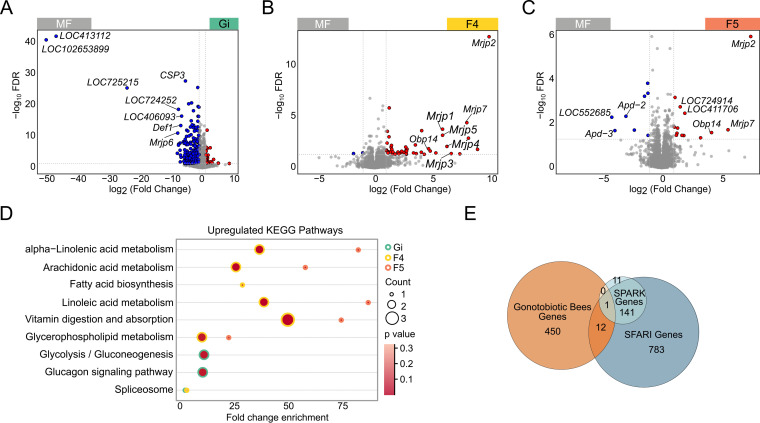
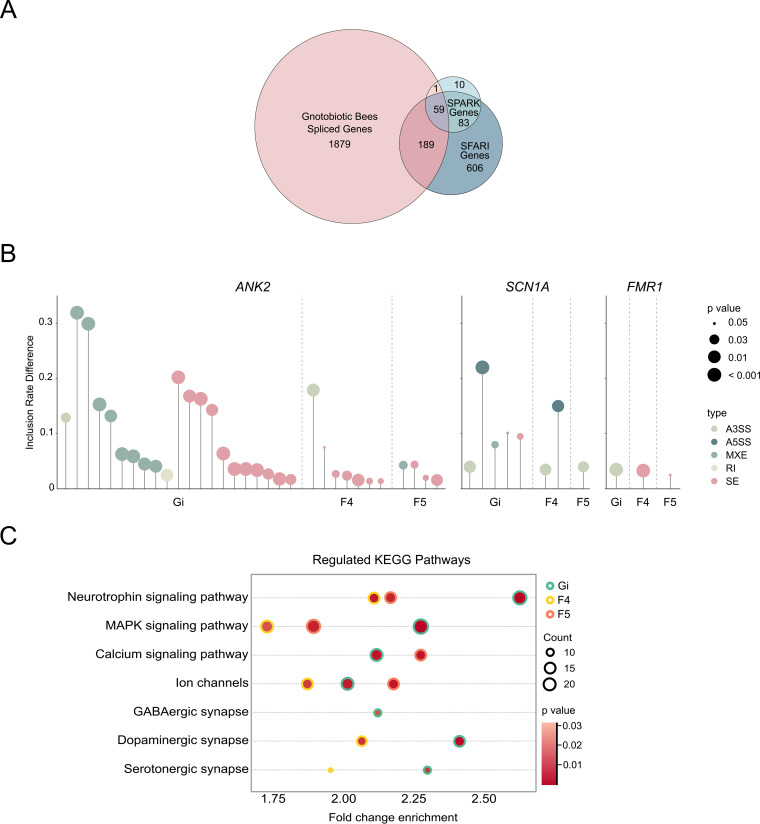
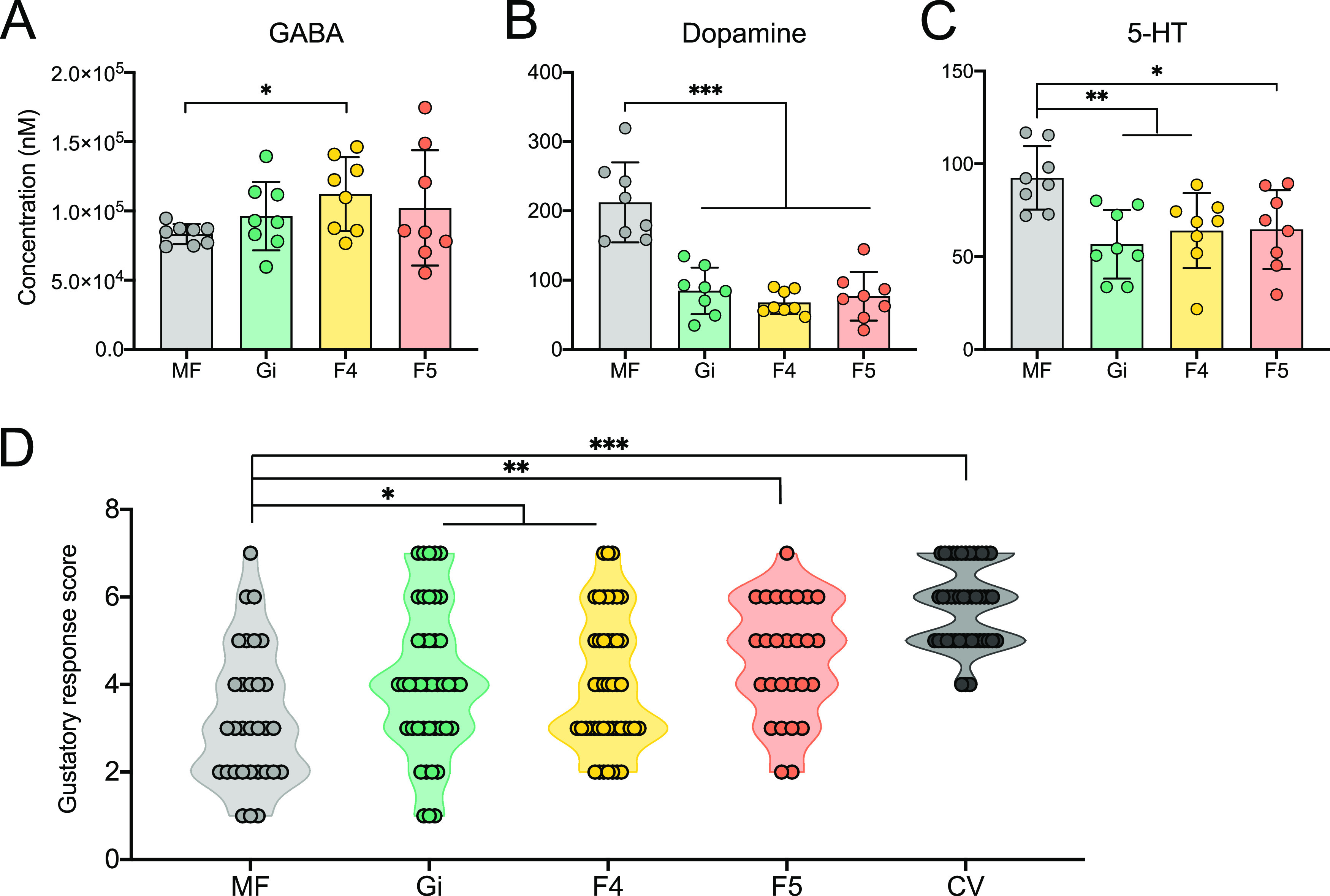
The honeybee possesses a limited number of bacterial phylotypes that play essential roles in host metabolism, hormonal signaling, and feeding behavior. However, the contribution of individual gut members in shaping honeybee brain profiles remains unclear. By generating gnotobiotic bees which were mono-colonized by a single gut bacterium, we revealed that different species regulated specific modules of metabolites in the hemolymph. Circulating metabolites involved in carbohydrate and glycerophospholipid metabolism pathways were mostly regulated by Gilliamella, while Lactobacillus Firm4 and Firm5 mainly altered amino acid metabolism pathways. We then analyzed the brain transcriptomes of bees mono-colonized with these three bacteria. These showed distinctive gene expression profiles, and genes related to olfactory functions and labor division were upregulated by Lactobacillus. Interestingly, differentially spliced genes in the brains of gnotobiotic bees largely overlapped with those of bees unresponsive to social stimuli. The differentially spliced genes were enriched in pathways involved in neural development and synaptic transmission. We showed that gut bacteria altered neurotransmitter levels in the brain. In particular, dopamine and serotonin, which show inhibitory effects on the sensory sensitivity of bees, were downregulated in bacteria-colonized bees. The proboscis extension response showed that a normal gut microbiota is essential for the taste-related behavior of honeybees, suggesting the contribution of potential interactions among different gut species to the host's physiology. Our findings provide fundamental insights into the diverse functions of gut bacteria which likely contribute to honeybee neurological processes. IMPORTANCE The honeybee possesses a simple and host-restricted gut community that contributes to the metabolic health of its host, while the effects of bacterial symbionts on host neural functions remain elusive. We found that the colonization of specific bee gut bacteria regulates distinct circulating metabolites enriched in carbohydrate, amino acid, and glycerophospholipid metabolic pathways. The brains of bees colonized with different gut members display distinct transcriptomic profiles of genes crucial for bee behaviors and division of labor. Alternative splicing of genes related to disordered bee behaviors is also mediated. The presence of gut bacteria promotes sucrose sensitivity with major neurotransmitters being regulated in the brain. Our findings demonstrate how individual bee gut species affect host behaviors, highlighting the gut-brain connections important for honeybee neurobiological and physiological states.
Keywords: Apis mellifera; behavior; gut-brain axis; metabolism; microbiome.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Raine NE, Ings TC, Dornhaus A, Selah N, Chittka L. 2006. Adaptation, genetic drift, pleiotropy, and history in the evolution of bee foraging behavior. Adv Stud Behav 36:305–354. doi: 10.1016/S0065-3454(06)36007-X. - DOI
-
- Scheiner R, Abramson CI, Brodschneider R, Crailsheim K, Farina WM, Fuchs S, Grünewald B, Hahshold S, Karrer M, Koeniger G, Koeniger N, Menzel R, Mujagic S, Radspieler G, Schmickl T, Schneider C, Siegel AJ, Szopek M, Thenius R. 2013. Standard methods for behavioural studies of Apis mellifera. J Apicult Res 52:1–58. doi: 10.3896/IBRA.1.52.4.04. - DOI
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
