

PYL1- and PYL8-like ABA Receptors of Nicotiana benthamiana Play a Key Role in ABA Response in Seed and Vegetative Tissue
- PMID: 35269417
- PMCID: PMC8909036
- DOI: 10.3390/cells11050795
PYL1- and PYL8-like ABA Receptors of Nicotiana benthamiana Play a Key Role in ABA Response in Seed and Vegetative Tissue
Abstract
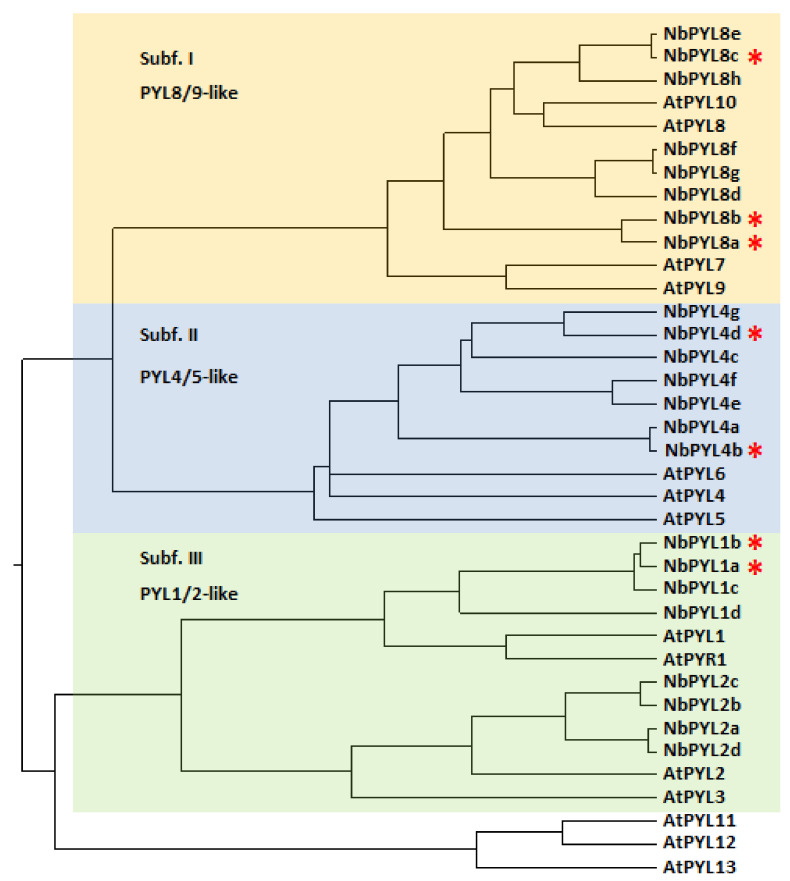
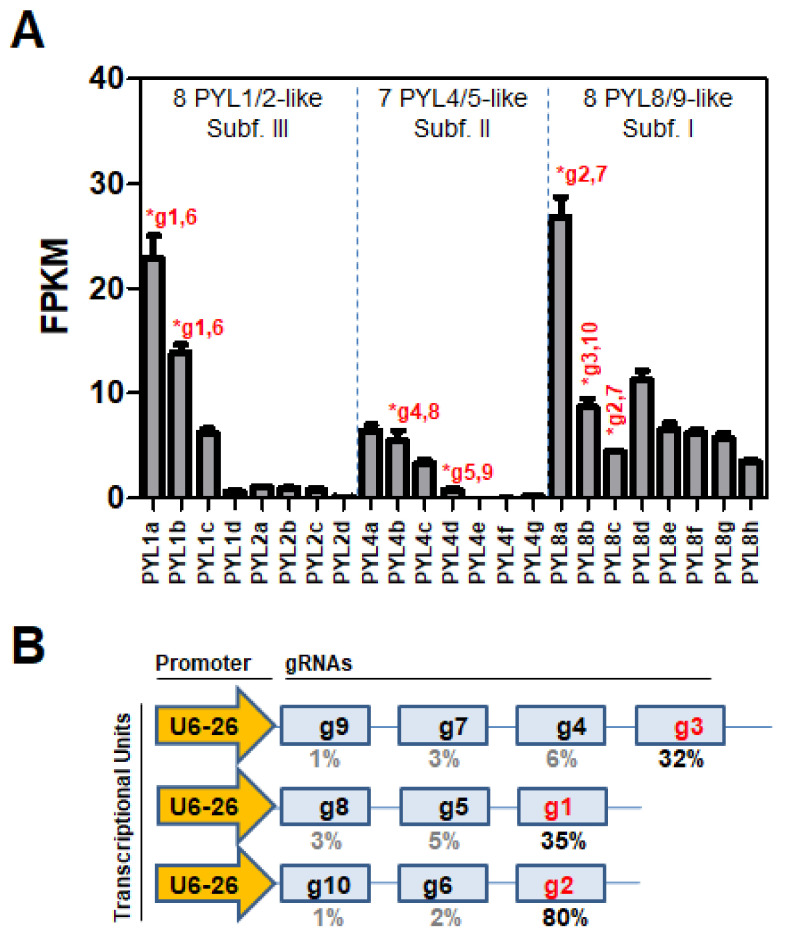
To face the challenges of climate change and sustainable food production, it is essential to develop crop genome editing techniques to pinpoint key genes involved in abiotic stress signaling. The identification of those prevailing abscisic acid (ABA) receptors that mediate plant-environment interactions is quite challenging in polyploid plants because of the high number of genes in the PYR/PYL/RCAR ABA receptor family. Nicotiana benthamiana is a biotechnological crop amenable to genome editing, and given the importance of ABA signaling in coping with drought stress, we initiated the analysis of its 23-member family of ABA receptors through multiplex CRISPR/Cas9-mediated editing. We generated several high-order mutants impaired in NbPYL1-like and NbPYL8-like receptors, which showed certain insensitivity to ABA for inhibition of seedling establishment, growth, and development of shoot and lateral roots as well as reduced sensitivity to the PYL1-agonist cyanabactin (CB). However, in these high-order mutants, regulation of transpiration was not affected and was responsive to ABA treatment. This reveals a robust and redundant control of transpiration in this allotetraploid plant that probably reflects its origin from the extreme habitat of central Australia.
Keywords: ABA receptors; ABA sensitivity; CRISPR/Cas9; Nicotiana benthamiana; biotechnological crop; drought; extremophile; gene editing; multiplex mutations; tetraploid.
Conflict of interest statement
Kacper P. Kaminski and Nikolai V. Ivanov are employees of Philip Morris International. The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures





Similar articles
-
Arabidopsis PYR/PYL/RCAR receptors play a major role in quantitative regulation of stomatal aperture and transcriptional response to abscisic acid.Plant Cell. 2012 Jun;24(6):2483-96. doi: 10.1105/tpc.112.098574. Epub 2012 Jun 26. Plant Cell. 2012. PMID: 22739828 Free PMC article.
-
Basal ABA signaling balances transpiration and photosynthesis.Physiol Plant. 2024 Sep-Oct;176(5):e14494. doi: 10.1111/ppl.14494. Physiol Plant. 2024. PMID: 39210540
-
Genome-wide identification and characterization of ABA receptor PYL/RCAR gene family reveals evolution and roles in drought stress in Nicotiana tabacum.BMC Genomics. 2019 Jul 11;20(1):575. doi: 10.1186/s12864-019-5839-2. BMC Genomics. 2019. PMID: 31296158 Free PMC article.
-
Arabidopsis Duodecuple Mutant of PYL ABA Receptors Reveals PYL Repression of ABA-Independent SnRK2 Activity.Cell Rep. 2018 Jun 12;23(11):3340-3351.e5. doi: 10.1016/j.celrep.2018.05.044. Cell Rep. 2018. PMID: 29898403 Free PMC article.
-
Structural insights into PYR/PYL/RCAR ABA receptors and PP2Cs.Plant Sci. 2012 Jan;182:3-11. doi: 10.1016/j.plantsci.2010.11.014. Epub 2010 Dec 7. Plant Sci. 2012. PMID: 22118610 Review.
Cited by
-
Transcriptional deregulation of stress-growth balance in Nicotiana benthamiana biofactories producing insect sex pheromones.Front Plant Sci. 2022 Oct 26;13:941338. doi: 10.3389/fpls.2022.941338. eCollection 2022. Front Plant Sci. 2022. PMID: 36388501 Free PMC article.
-
StSN2 enhances tuber formation in potato via upregulating of the ABA signaling pathway.Front Plant Sci. 2025 May 12;16:1566237. doi: 10.3389/fpls.2025.1566237. eCollection 2025. Front Plant Sci. 2025. PMID: 40420855 Free PMC article.
-
Genome-wide identification, phylogenetic, structural and functional evolution of the core components of ABA signaling in plant species: a focus on rice.Planta. 2024 Jul 22;260(3):58. doi: 10.1007/s00425-024-04475-2. Planta. 2024. PMID: 39039384
