

A Human In Vitro Model to Study Adenoviral Receptors and Virus Cell Interactions
- PMID: 35269463
- PMCID: PMC8909167
- DOI: 10.3390/cells11050841
A Human In Vitro Model to Study Adenoviral Receptors and Virus Cell Interactions
Abstract
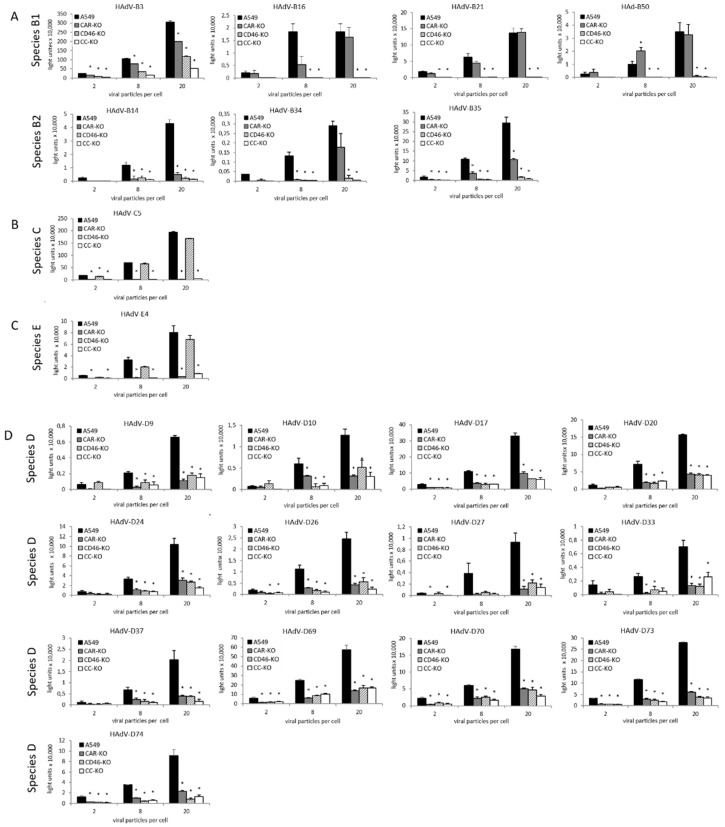
To develop adenoviral cell- or tissue-specific gene delivery, understanding of the infection mechanisms of adenoviruses is crucial. Several adenoviral attachment proteins such as CD46, CAR and sialic acid have been identified and studied. However, most receptor studies were performed on non-human cells. Combining our reporter gene-tagged adenovirus library with an in vitro human gene knockout model, we performed a systematic analysis of receptor usage comparing different adenoviruses side-by-side. The CRISPR/Cas9 system was used to knockout CD46 and CAR in the human lung epithelial carcinoma cell line A549. Knockout cells were infected with 22 luciferase-expressing adenoviruses derived from adenovirus species B, C, D and E. HAdV-B16, -B21 and -B50 from species B1 as well as HAdV-B34 and -B35 were found to be CD46-dependent. HAdV-C5 and HAdV-E4 from species E were found to be CAR-dependent. Regarding cell entry of HAdV-B3 and -B14 and all species D viruses, both CAR and CD46 play a role, and here, other receptors or attachment structures may also be important since transductions were reduced but not completely inhibited. The established human knockout cell model enables the identification of the most applicable adenovirus types for gene therapy and to further understand adenovirus infection biology.
Keywords: CAR; CD46; CD46-knockout cell line; CRISPR; adenovirus; human; knockout cell line; receptor; tight-junction-knockout cell line; virus.
Conflict of interest statement
The authors declare no conflict of interest.
Figures





References
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
