

The current toolbox for APOBEC drug discovery
- PMID: 35272863
- PMCID: PMC9018551
- DOI: 10.1016/j.tips.2022.02.007
The current toolbox for APOBEC drug discovery
Abstract
Mutational processes driving genome evolution and heterogeneity contribute to immune evasion and therapy resistance in viral infections and cancer. APOBEC3 (A3) enzymes promote such mutations by catalyzing the deamination of cytosines to uracils in single-stranded DNA. Chemical inhibition of A3 enzymes may yield an antimutation therapeutic strategy to improve the durability of current drug therapies that are prone to resistance mutations. A3 small-molecule drug discovery efforts to date have been restricted to a single high-throughput biochemical activity assay; however, the arsenal of discovery assays has significantly expanded in recent years. The assays used to study A3 enzymes are reviewed here with an eye towards their potential for small-molecule discovery efforts.
Keywords: APOBEC; DNA deaminase inhibitors; chemical probes; drug discovery; screening.
Copyright © 2022 The Authors. Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
Declaration of interests R.S.H. and D.A.H. were cofounders of ApoGen Biotechnologies, Inc., which closed operations in April 2021. The other authors declare no conflicts of interest.
Figures
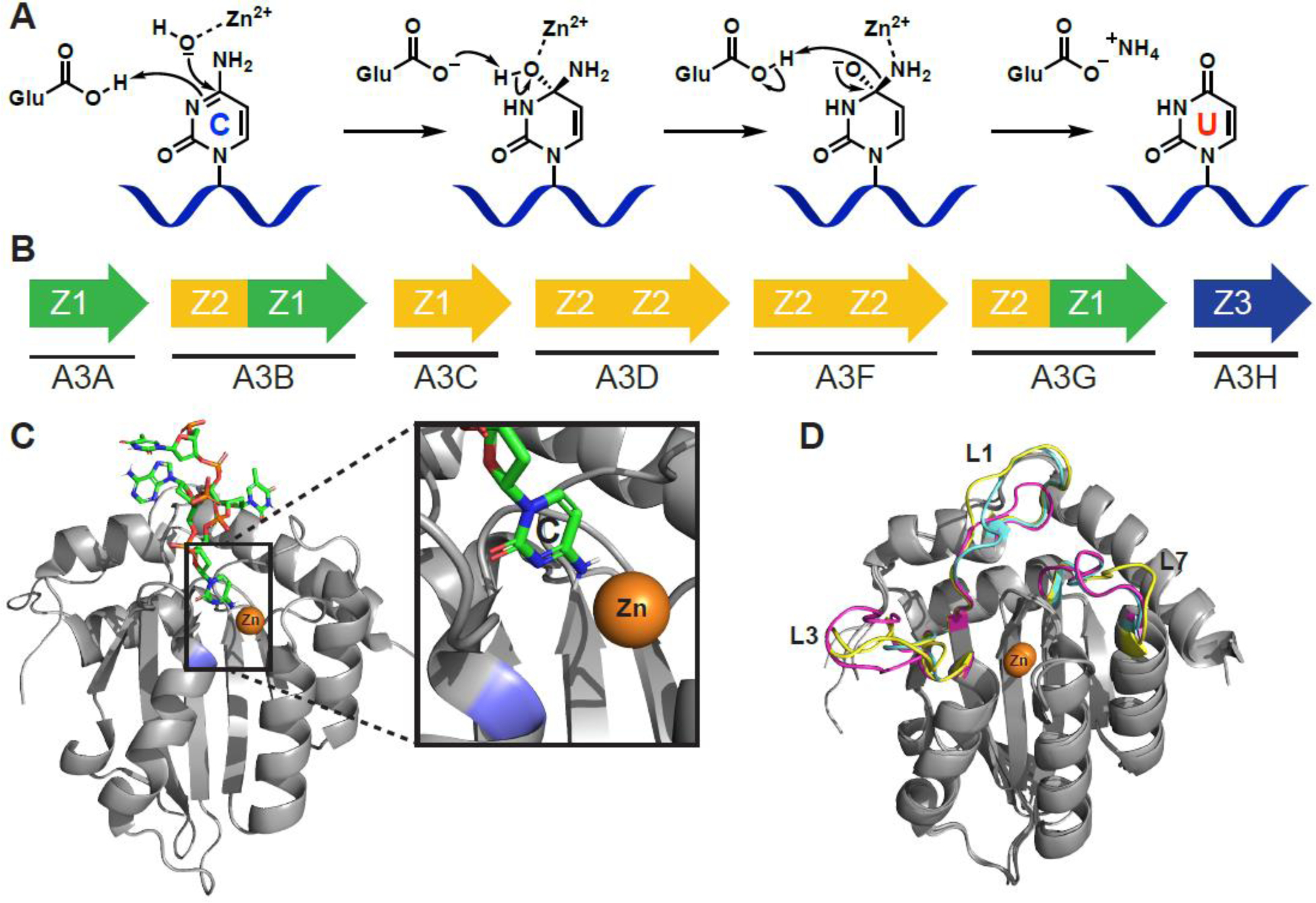
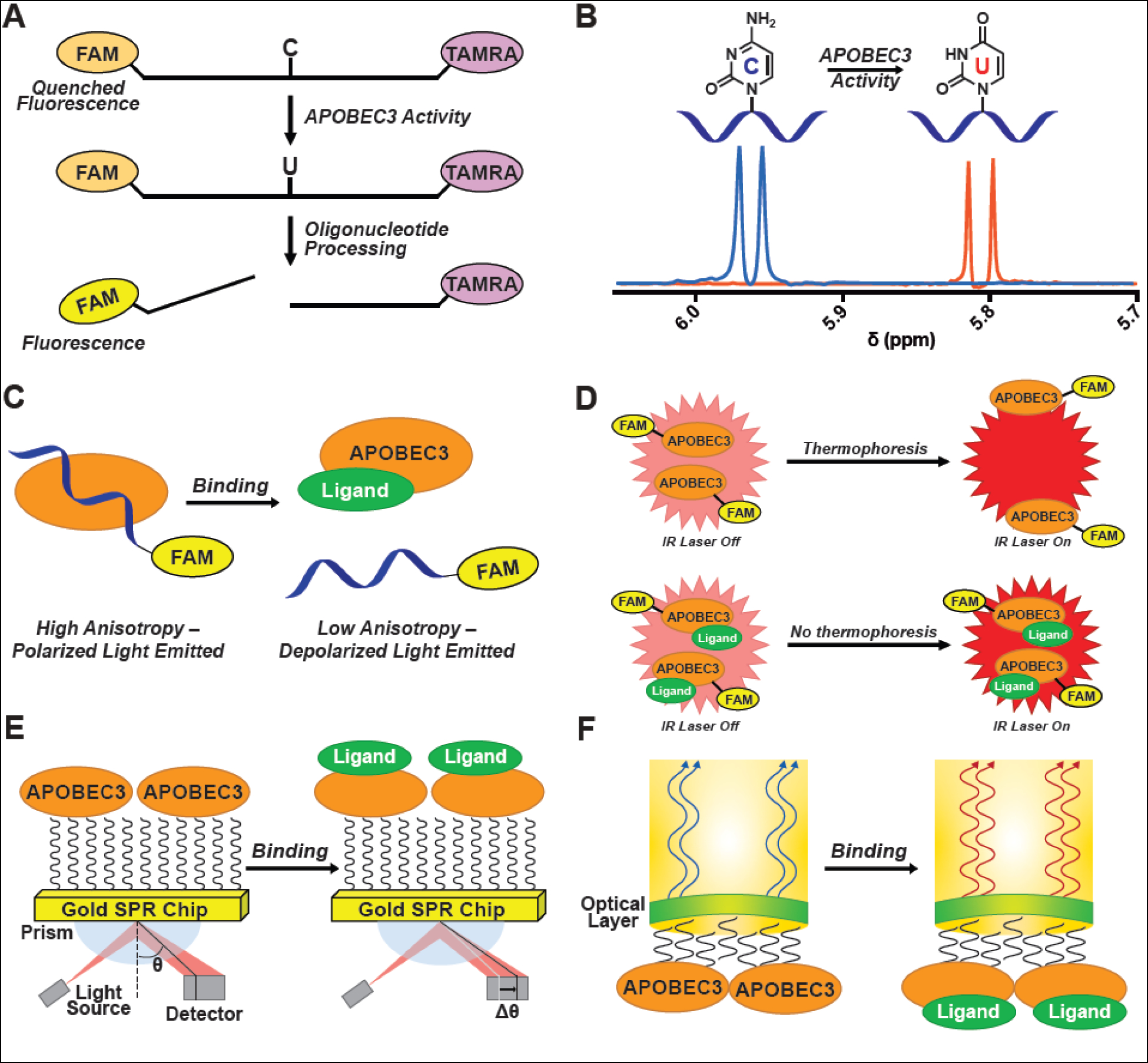
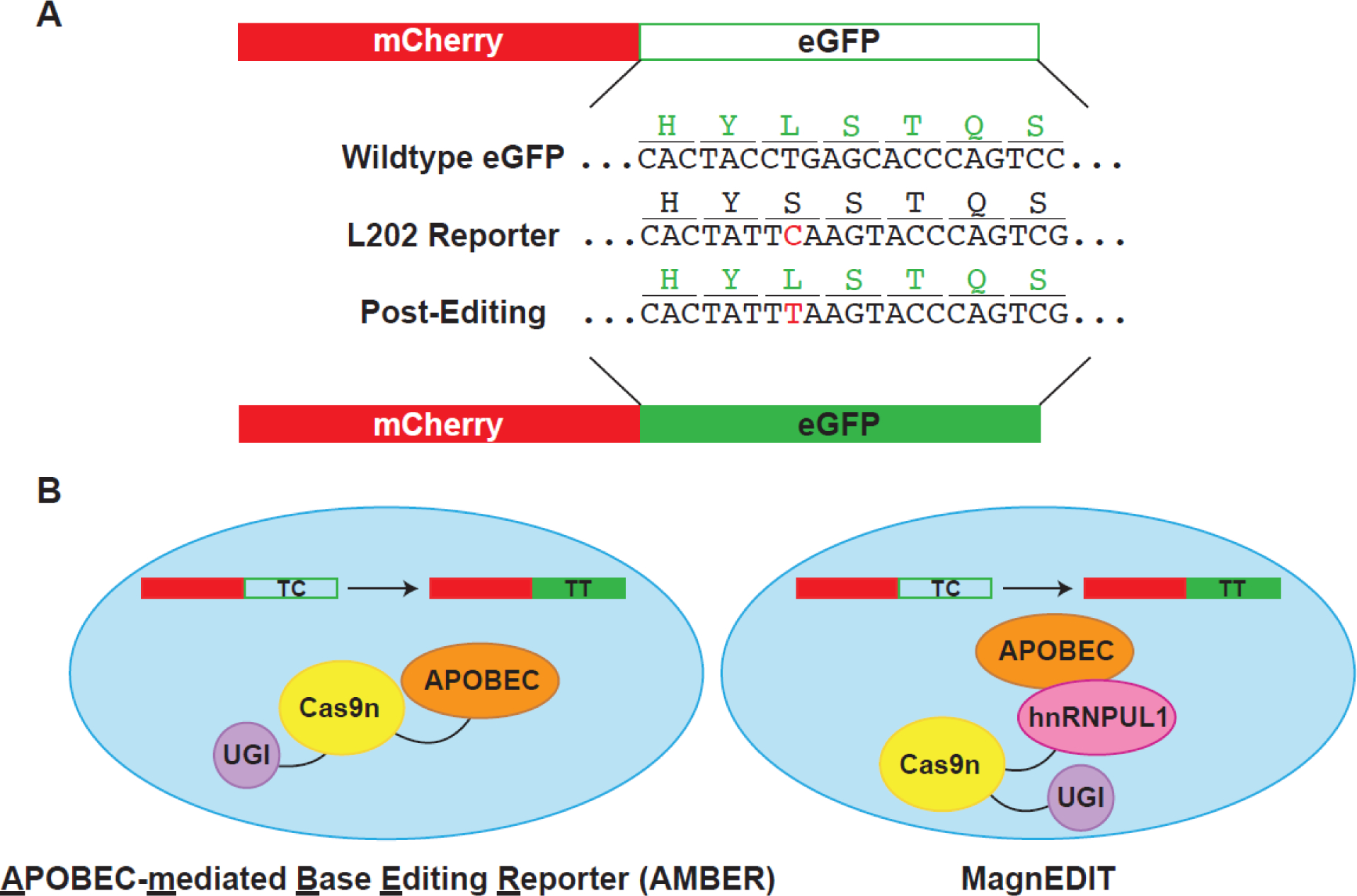
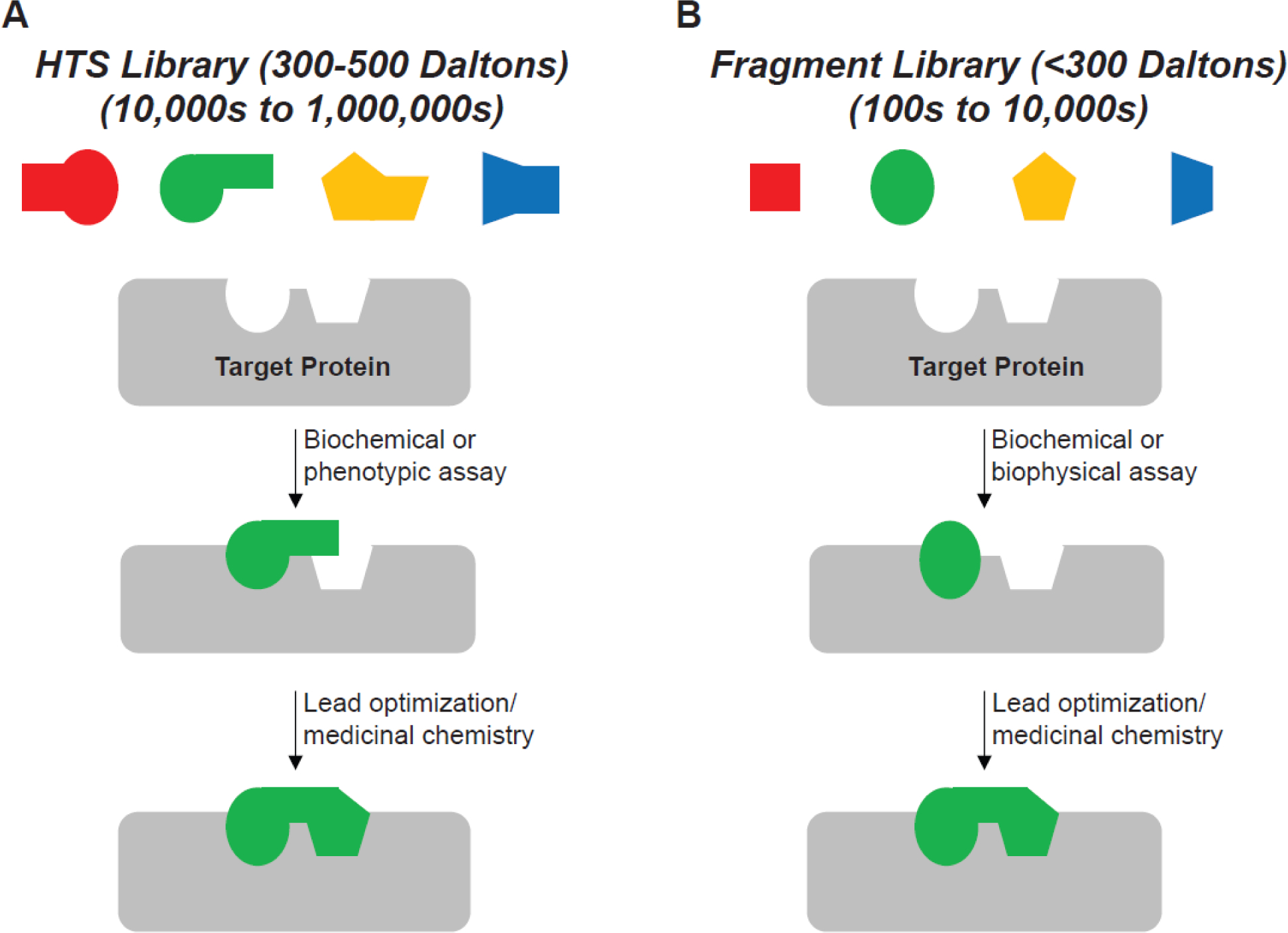
References
-
- Betts L, et al. (1994) Cytidine deaminase. The 2.3 Å crystal structure of an enzyme: transition-state analog complex. J. Mol. Biol. 235, 635–656 - PubMed
-
- Johansson E, et al. (2002) Crystal structure of the tetrameric cytidine deaminase from Bacillus subtilis at 2.0 Å resolution. Biochemistry 41, 2563–2570 - PubMed
-
- Yao L, et al. (2005) A molecular dynamics exploration of the catalytic mechanism of yeast cytosine deaminase. J. Phys. Chem. B 109, 7500–7510 - PubMed
-
- Manta B, et al. (2014) Reaction Mechanism of Zinc-Dependent Cytosine Deaminase from Escherichia coli: A Quantum-Chemical Study. J. Phys. Chem. B 118, 5644–5652 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
