

Metabolic and Epigenetic Regulation by Estrogen in Adipocytes
- PMID: 35273571
- PMCID: PMC8901598
- DOI: 10.3389/fendo.2022.828780
Metabolic and Epigenetic Regulation by Estrogen in Adipocytes
Abstract
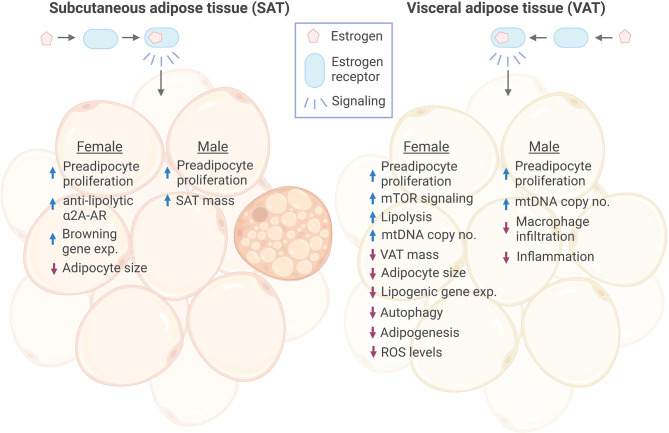
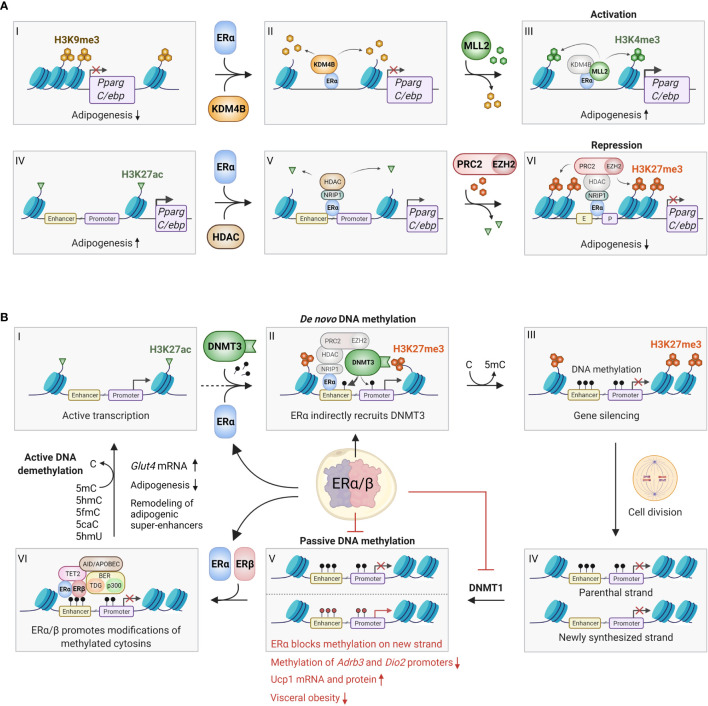
Sex hormones contribute to differences between males and females in body fat distribution and associated disease risk. Higher concentrations of estrogens are associated with a more gynoid body shape and with more fat storage on hips and thighs rather than in visceral depots. Estrogen-mediated protection against visceral adiposity is shown in post-menopausal women with lower levels of estrogens and the reduction in central body fat observed after treatment with hormone-replacement therapy. Estrogen exerts its physiological effects via the estrogen receptors (ERα, ERβ and GPR30) in target cells, including adipocytes. Studies in mice indicate that estrogen protects against adipose inflammation and fibrosis also before the onset of obesity. The mechanisms involved in estrogen-dependent body fat distribution are incompletely understood, but involve, e.g., increased mTOR signaling and suppression of autophagy and adipogenesis/lipid storage. Estrogen plays a key role in epigenetic regulation of adipogenic genes by interacting with enzymes that remodel DNA methylation and histone tail post-translational modifications. However, more studies are needed to map the differential epigenetic effects of ER in different adipocyte subtypes, including those in subcutaneous and visceral adipose tissues. We here review recent discoveries of ER-mediated transcriptional and epigenetic regulation in adipocytes, which may explain sexual dimorphisms in body fat distribution and obesity-related disease risk.
Keywords: adipocyte; epigenetic; estrogen; sexual dimorphism; steroids.
Copyright © 2022 Bjune, Strømland, Jersin, Mellgren and Dankel.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous