

d-Serine controls epidermal vesicle release via NMDA receptor, allowing tissue migration during the metamorphosis of the chordate Ciona
- PMID: 35275721
- PMCID: PMC8916719
- DOI: 10.1126/sciadv.abn3264
d-Serine controls epidermal vesicle release via NMDA receptor, allowing tissue migration during the metamorphosis of the chordate Ciona
Abstract
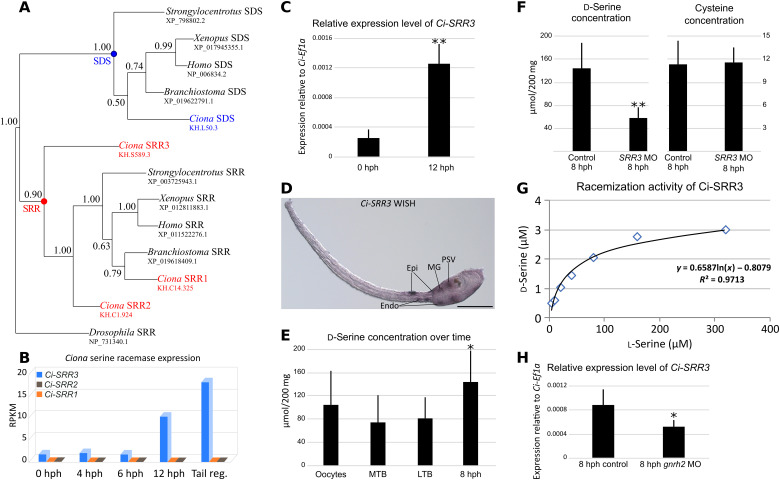
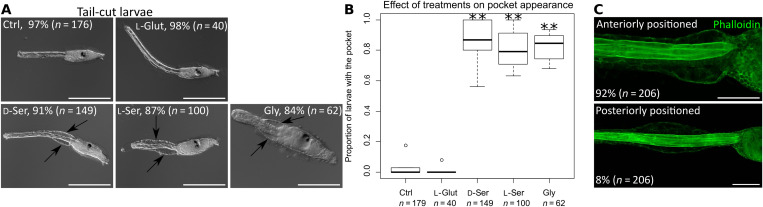
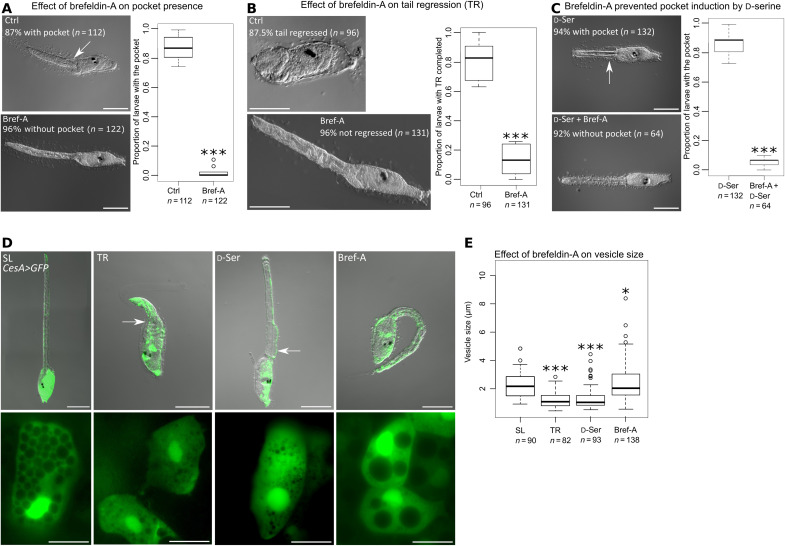
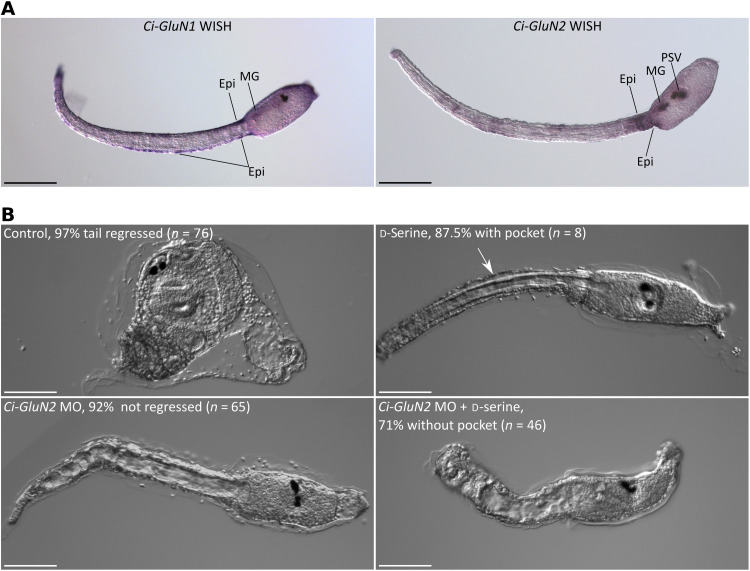
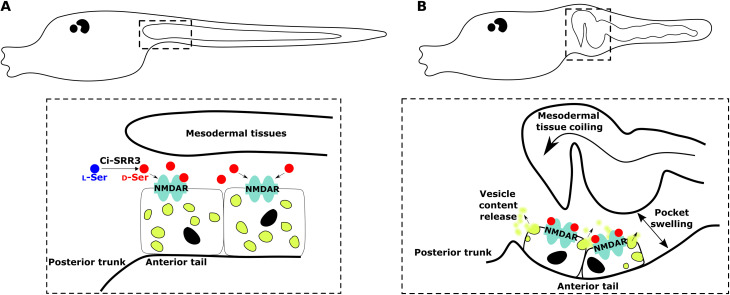
d-Serine, a free amino acid synthesized by serine racemase, is a coagonist of N-methyl-d-aspartate-type glutamate receptor (NMDAR). d-Serine in the mammalian central nervous system modulates glutamatergic transmission. Functions of d-serine in mammalian peripheral tissues such as skin have also been described. However, d-serine's functions in nonmammals are unclear. Here, we characterized d-serine-dependent vesicle release from the epidermis during metamorphosis of the tunicate Ciona. d-Serine leads to the formation of a pocket that facilitates the arrival of migrating tissue during tail regression. NMDAR is the receptor of d-serine in the formation of the epidermal pocket. The epidermal pocket is formed by the release of epidermal vesicles' content mediated by d-serine/NMDAR. This mechanism is similar to observations of keratinocyte vesicle exocytosis in mammalian skin. Our findings provide a better understanding of the maintenance of epidermal homeostasis in animals and contribute to further evolutionary perspectives of d-amino acid function among metazoans.
Figures

Similar articles
-
Extrinsic apoptosis participates to tail regression during the metamorphosis of the chordate Ciona.Sci Rep. 2024 Mar 8;14(1):5729. doi: 10.1038/s41598-023-48411-y. Sci Rep. 2024. PMID: 38459045 Free PMC article.
-
AMPA receptor-dependent, light-evoked D-serine release acts on retinal ganglion cell NMDA receptors.J Neurophysiol. 2012 Aug;108(4):1044-51. doi: 10.1152/jn.00264.2012. Epub 2012 May 16. J Neurophysiol. 2012. PMID: 22592312 Free PMC article.
-
Localization of serine racemase and its role in the skin.J Invest Dermatol. 2014 Jun;134(6):1618-1626. doi: 10.1038/jid.2014.22. Epub 2014 Jan 17. J Invest Dermatol. 2014. PMID: 24441099 Free PMC article.
-
D-serine regulation of NMDA receptor activity.Sci STKE. 2006 Oct 10;2006(356):pe41. doi: 10.1126/stke.3562006pe41. Sci STKE. 2006. PMID: 17033043 Review.
-
Analysis of free D-serine in mammals and its biological relevance.J Chromatogr B Analyt Technol Biomed Life Sci. 2011 Nov 1;879(29):3169-83. doi: 10.1016/j.jchromb.2011.08.030. Epub 2011 Aug 30. J Chromatogr B Analyt Technol Biomed Life Sci. 2011. PMID: 21992750 Review.
Cited by
-
Extrinsic apoptosis participates to tail regression during the metamorphosis of the chordate Ciona.Sci Rep. 2024 Mar 8;14(1):5729. doi: 10.1038/s41598-023-48411-y. Sci Rep. 2024. PMID: 38459045 Free PMC article.
-
Serine metabolism in aging and age-related diseases.Geroscience. 2025 Feb;47(1):611-630. doi: 10.1007/s11357-024-01444-1. Epub 2024 Nov 25. Geroscience. 2025. PMID: 39585647 Free PMC article. Review.
-
Chiral derivatization-enabled discrimination and on-tissue detection of proteinogenic amino acids by ion mobility mass spectrometry.Chem Sci. 2022 Nov 21;13(47):14114-14123. doi: 10.1039/d2sc03604e. eCollection 2022 Dec 7. Chem Sci. 2022. PMID: 36540812 Free PMC article.
References
-
- Hashimoto A., Nishikawa T., Hayashi T., Fujii N., Harada K., Oka T., Takahashi K., The presence of free D-serine in rat brain. FEBS Lett. 296, 33–36 (1992). - PubMed
-
- Matsui T., Sekiguchi M., Hashimoto A., Tomita U., Nishikawa T., Wada K., Functional comparison of D-serine and glycine in rodents: The effect on cloned NMDA receptors and the extracellular concentration. J. Neurochem. 65, 454–458 (1995). - PubMed
-
- Priestley T., Laughton P., Myers J., Le Bourdellés B., Kerby J., Whiting P. J., Pharmacological properties of recombinant human N-methyl-D-aspartate receptors comprising NR1a/NR2A and NR1a/NR2B subunit assemblies expressed in permanently transfected mouse fibroblast cells. Mol. Pharmacol. 48, 841–848 (1995). - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
