

Lamin B1 deletion in myeloid neoplasms causes nuclear anomaly and altered hematopoietic stem cell function
- PMID: 35278369
- PMCID: PMC9018112
- DOI: 10.1016/j.stem.2022.02.010
Lamin B1 deletion in myeloid neoplasms causes nuclear anomaly and altered hematopoietic stem cell function
Abstract
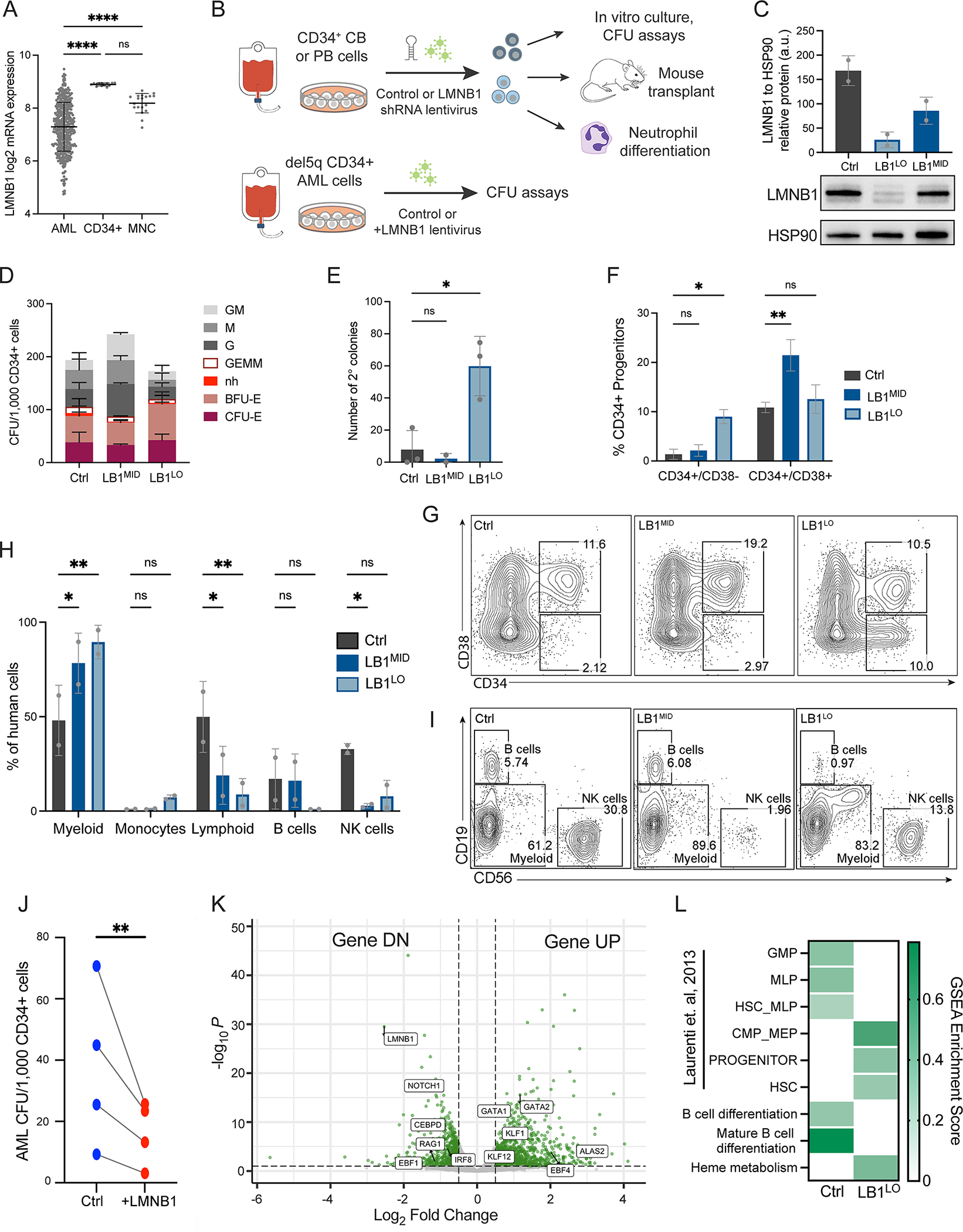
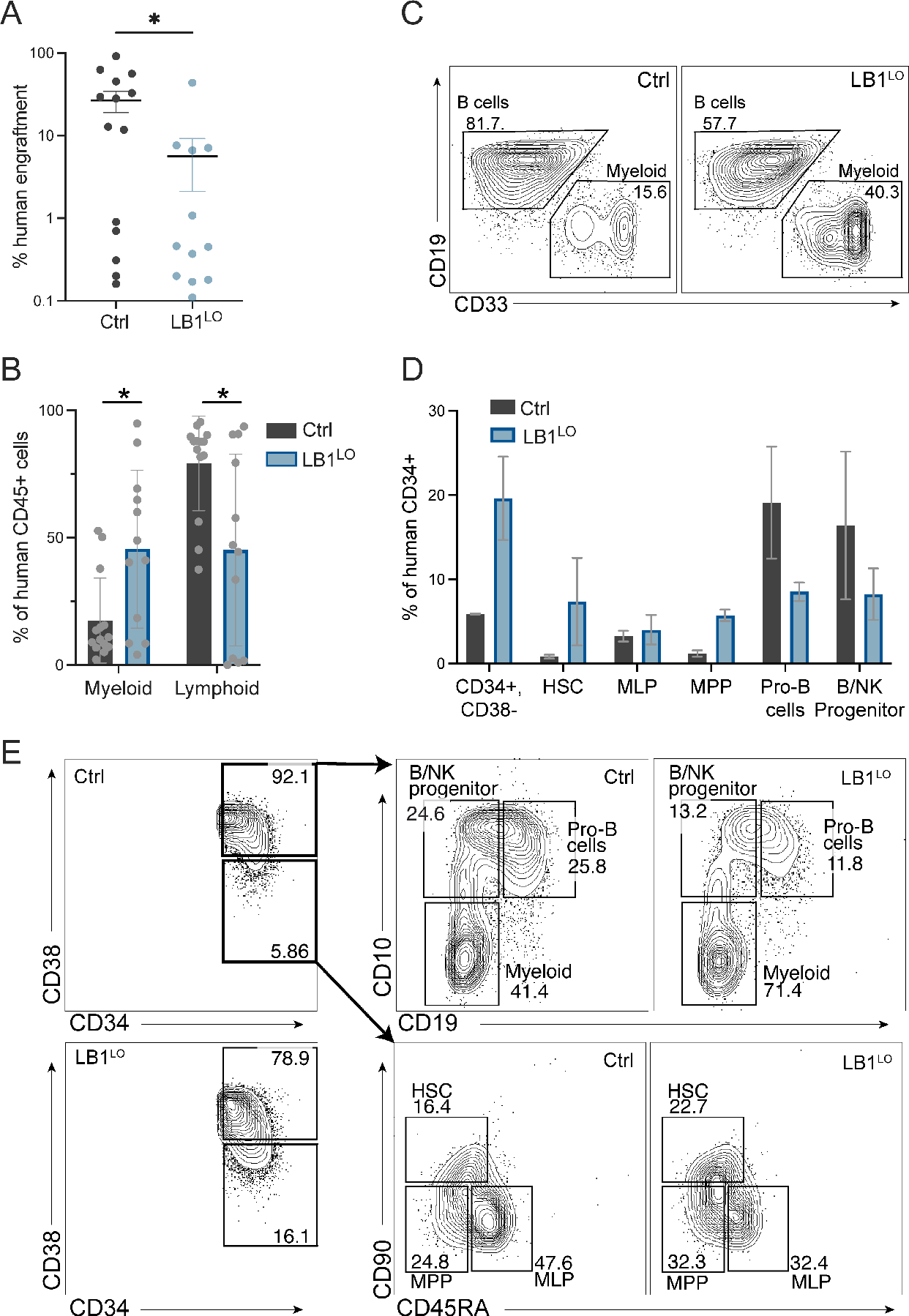
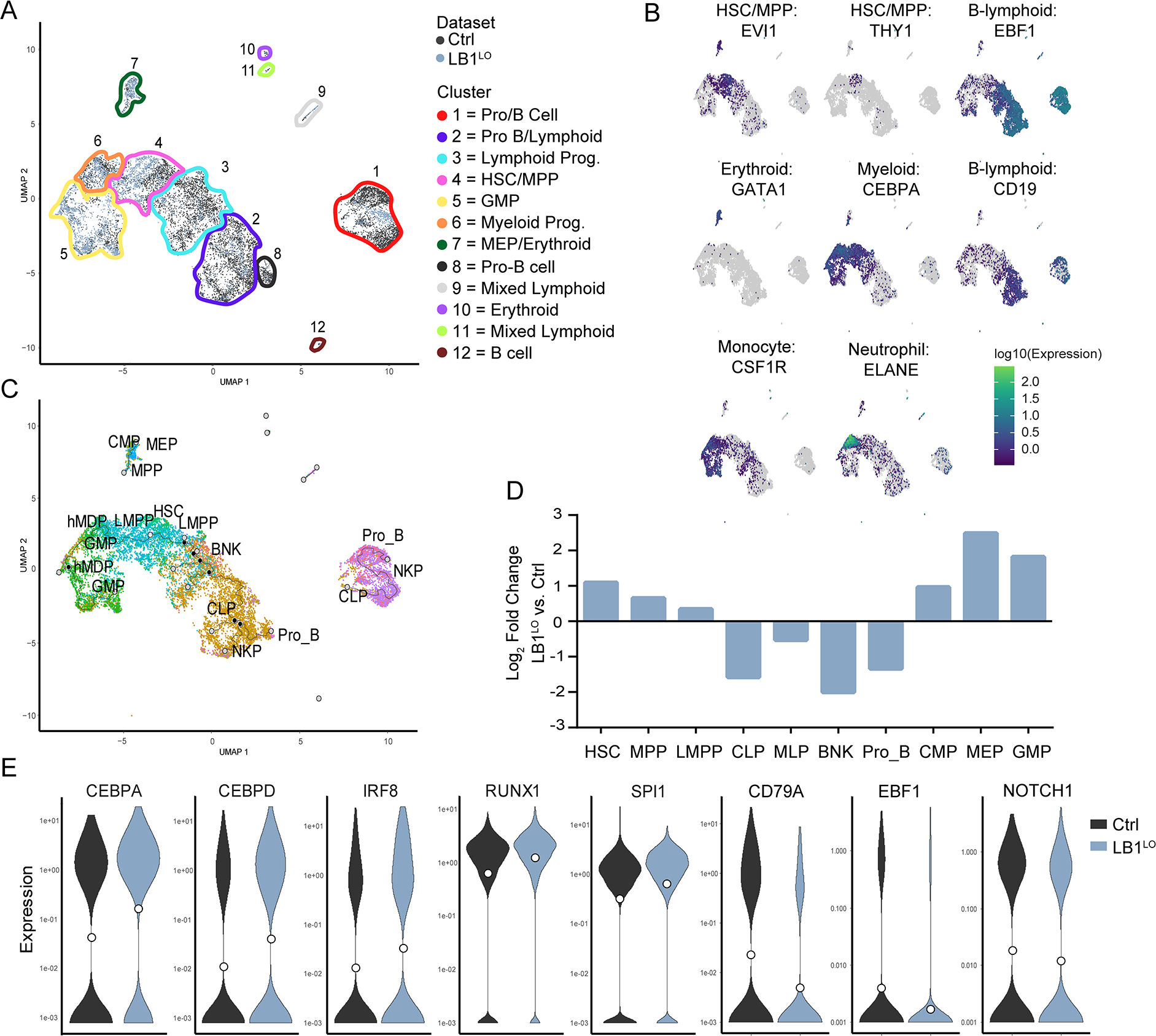
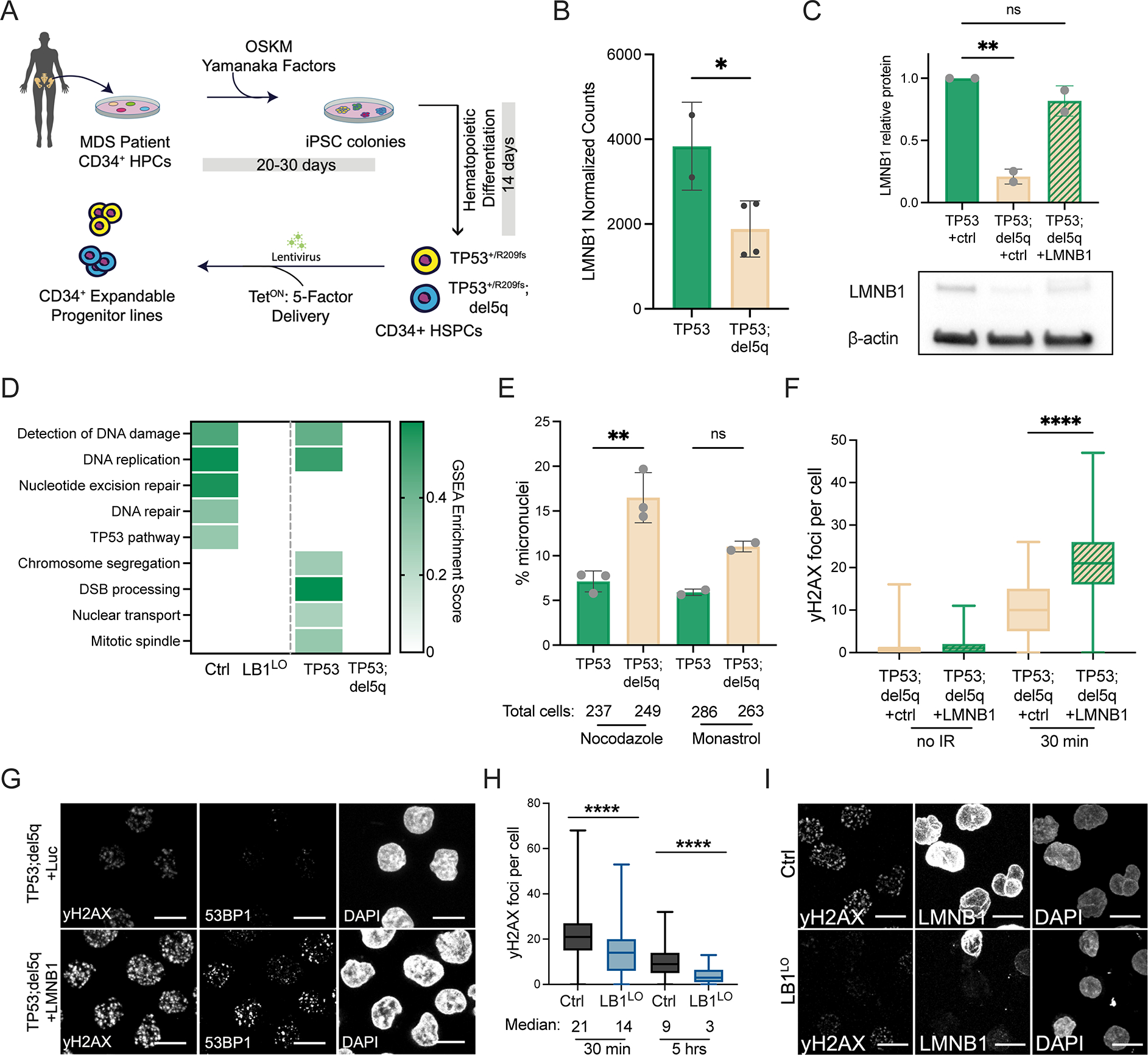
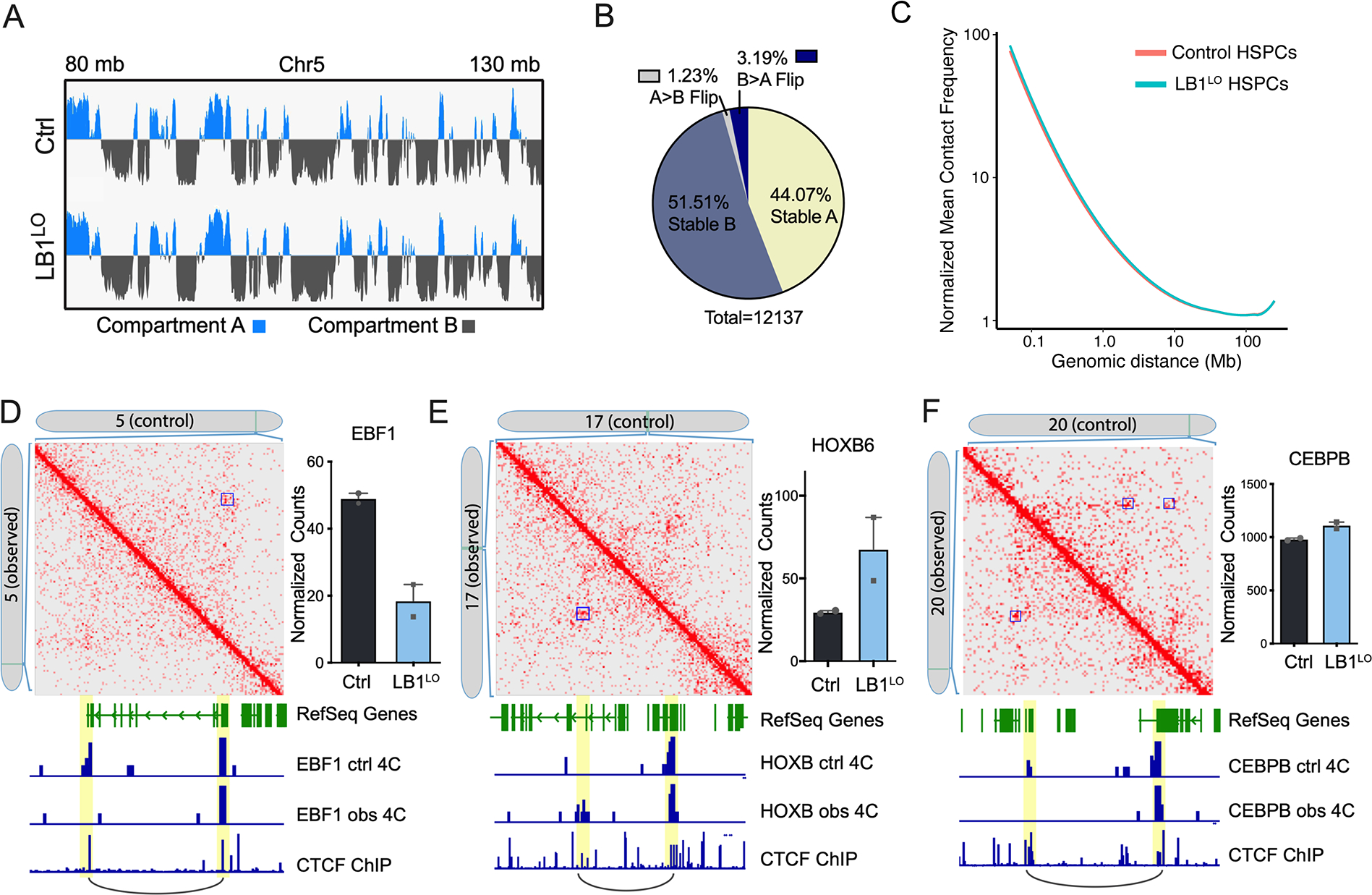
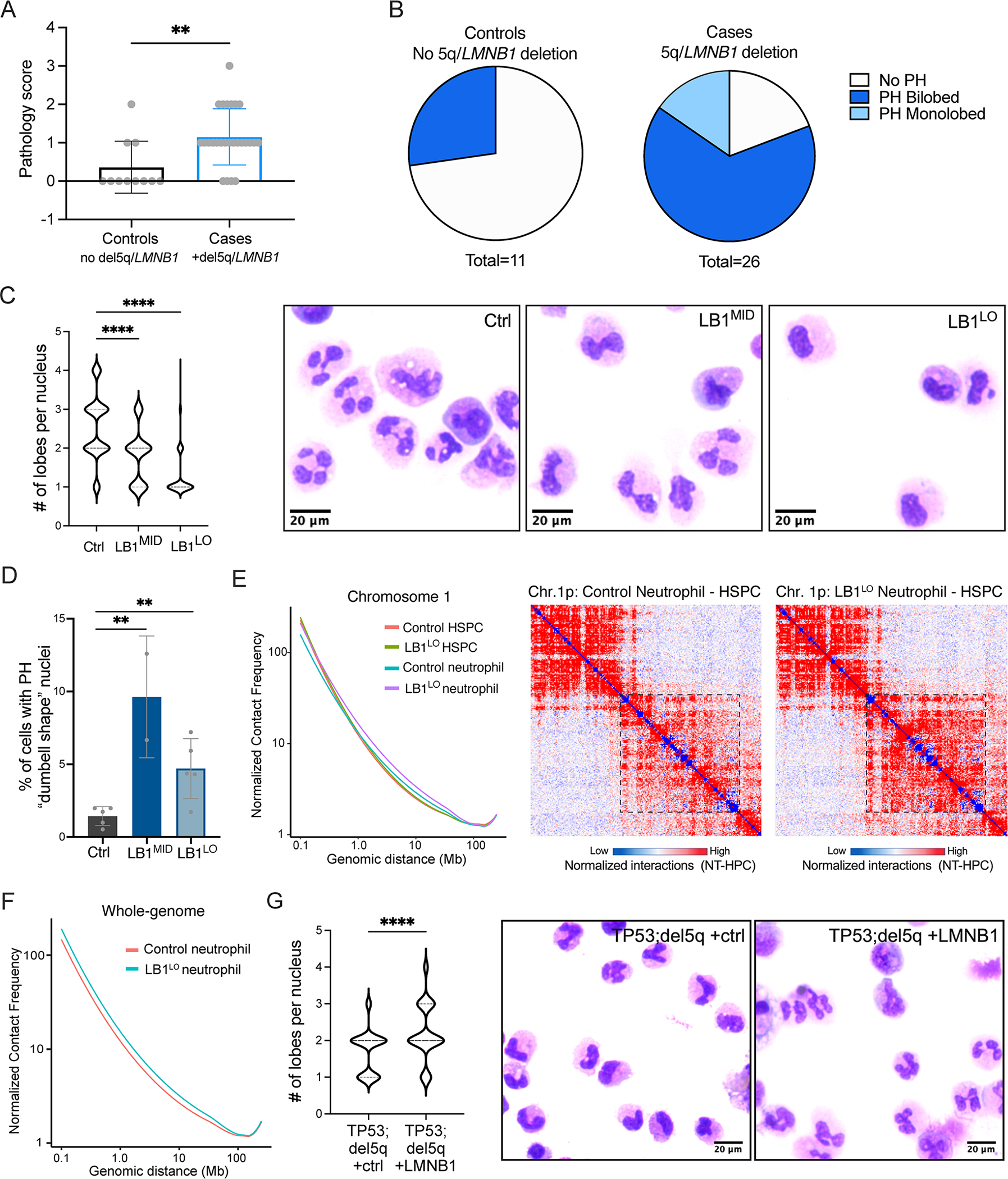
Abnormal nuclear morphology is a hallmark of malignant cells widely used in cancer diagnosis. Pelger-Huët anomaly (PHA) is a common abnormality of neutrophil nuclear morphology of unknown molecular etiology in myeloid neoplasms (MNs). We show that loss of nuclear lamin B1 (LMNB1) encoded on chromosome 5q, which is frequently deleted in MNs, induces defects in nuclear morphology and human hematopoietic stem cell (HSC) function associated with malignancy. LMNB1 deficiency alters genome organization inducing in vitro and in vivo expansion of HSCs, myeloid-biased differentiation with impaired lymphoid commitment, and genome instability due to defective DNA damage repair. Nuclear dysmorphology of neutrophils in patients with MNs is associated with 5q deletions spanning the LMNB1 locus, and lamin B1 loss is both necessary and sufficient to cause PHA in normal and 5q-deleted neutrophils. LMNB1 loss thus causes acquired PHA and links abnormal nuclear morphology with HSCs and progenitor cell fate determination via genome organization.
Keywords: 3D genome; 5q deletions; genome instability; hematopoietic stem and progenitor cells; lineage determination; myeloid neoplasms; neutrophils; nuclear lamins; nuclear morphology.
Copyright © 2022 Elsevier Inc. All rights reserved.
Conflict of interest statement
Declaration of interests The authors declare no competing interests.
Figures
Comment in
-
MDS/AML with del5q: An acquired "laminopathy"?Cell Stem Cell. 2022 Apr 7;29(4):498-499. doi: 10.1016/j.stem.2022.03.008. Cell Stem Cell. 2022. PMID: 35395184 Free PMC article.
Similar articles
-
MDS/AML with del5q: An acquired "laminopathy"?Cell Stem Cell. 2022 Apr 7;29(4):498-499. doi: 10.1016/j.stem.2022.03.008. Cell Stem Cell. 2022. PMID: 35395184 Free PMC article.
-
[Nuclear abnormalities in Pelger-Huet anomaly; progress in blood cell morphology].Rinsho Byori. 2005 Jan;53(1):54-60. Rinsho Byori. 2005. PMID: 15724491 Review. Japanese.
-
Mutations in the gene encoding the lamin B receptor produce an altered nuclear morphology in granulocytes (Pelger-Huët anomaly).Nat Genet. 2002 Aug;31(4):410-4. doi: 10.1038/ng925. Epub 2002 Jul 15. Nat Genet. 2002. PMID: 12118250
-
An in vitro model for Pelger-Huët anomaly: stable knockdown of lamin B receptor in HL-60 cells.Nucleus. 2010 Nov-Dec;1(6):506-12. doi: 10.4161/nucl.1.6.13271. Epub 2010 Aug 6. Nucleus. 2010. PMID: 21327094 Free PMC article.
-
Understanding and recognizing the Pelger-Huët anomaly.Am J Clin Pathol. 2012 Mar;137(3):358-66. doi: 10.1309/AJCP3G8MDUXYSCID. Am J Clin Pathol. 2012. PMID: 22338047 Review.
Cited by
-
First evidence of coexistence of Pseudo Pelger Huet anomaly and balanced translocation in a two decades retrospectively exposed human subject.Sci Rep. 2025 Aug 11;15(1):29292. doi: 10.1038/s41598-025-14858-4. Sci Rep. 2025. PMID: 40790071 Free PMC article.
-
MDS/AML with del5q: An acquired "laminopathy"?Cell Stem Cell. 2022 Apr 7;29(4):498-499. doi: 10.1016/j.stem.2022.03.008. Cell Stem Cell. 2022. PMID: 35395184 Free PMC article.
-
Tuning between Nuclear Organization and Functionality in Health and Disease.Cells. 2023 Feb 23;12(5):706. doi: 10.3390/cells12050706. Cells. 2023. PMID: 36899842 Free PMC article. Review.
-
A generalizable Hi-C foundation model for chromatin architecture, single-cell and multi-omics analysis across species.bioRxiv [Preprint]. 2024 Dec 20:2024.12.16.628821. doi: 10.1101/2024.12.16.628821. bioRxiv. 2024. PMID: 39763871 Free PMC article. Preprint.
-
Immune cells adapt to confined environments in vivo to optimise nuclear plasticity for migration.EMBO Rep. 2025 Mar;26(5):1238-1268. doi: 10.1038/s44319-025-00381-0. Epub 2025 Feb 6. EMBO Rep. 2025. PMID: 39915297 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
