

Ion flux-independent NMDA receptor signaling
- PMID: 35278420
- PMCID: PMC9365119
- DOI: 10.1016/j.neuropharm.2022.109019
Ion flux-independent NMDA receptor signaling
Abstract
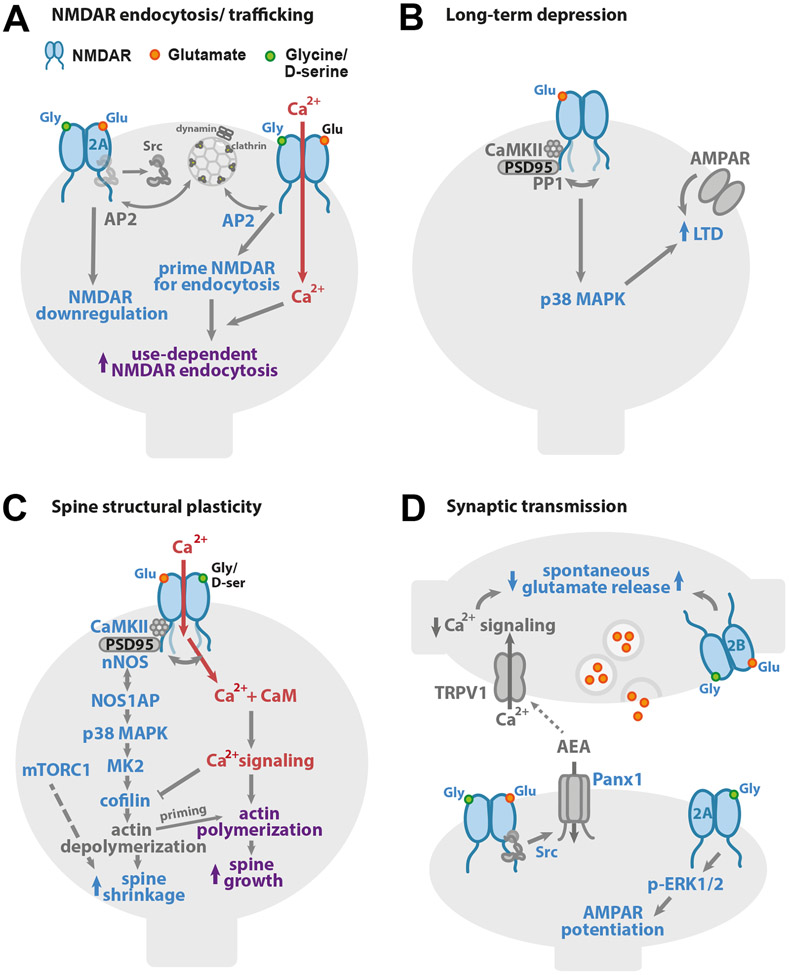
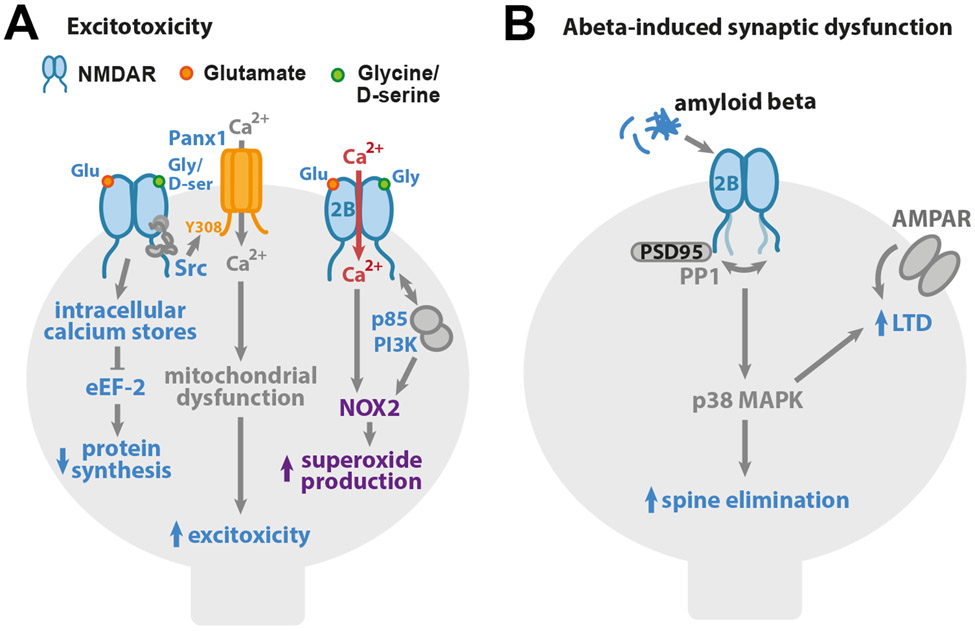
NMDA receptors play vital roles in a broad array of essential brain functions, from synaptic transmission and plasticity to learning and memory. Historically, the fundamental roles of NMDARs were attributed to their specialized properties of ion flux. More recently, it has become clear that NMDARs also signal in an ion flux-independent manner. Here, we review these non-ionotropic NMDAR signaling mechanisms that have been reported to contribute to a broad array of neuronal functions and dysfunctions including synaptic transmission and plasticity, cell death and survival, and synaptic alterations associated with neurological disorders.
Keywords: Alzheimer's disease; Dendritic spine; Ion flux-independent; LTD; LTP; NMDA receptor; Non-ionotropic; Schizophrenia.
Copyright © 2022 The Authors. Published by Elsevier Ltd.. All rights reserved.
Conflict of interest statement
Figures
References
-
- Aarts M, Liu Y, Liu L, Besshoh S, Arundine M, Gurd JW, Wang YT, Salter MW, and Tymianski M (2002). Treatment of ischemic brain damage by perturbing NMDA receptor- PSD-95 protein interactions. Science 298, 846–850. - PubMed
-
- Abrahamsson T, Chou CYC, Li SY, Mancino A, Costa RP, Brock JA, Nuro E, Buchanan KA, Elgar D, Blackman AV, et al. (2017). Differential Regulation of Evoked and Spontaneous Release by Presynaptic NMDA Receptors. Neuron 96, 839–855 e835. - PubMed
