

Key features of inhibitor binding to the human mitochondrial pyruvate carrier hetero-dimer
- PMID: 35278701
- PMCID: PMC8968063
- DOI: 10.1016/j.molmet.2022.101469
Key features of inhibitor binding to the human mitochondrial pyruvate carrier hetero-dimer
Abstract
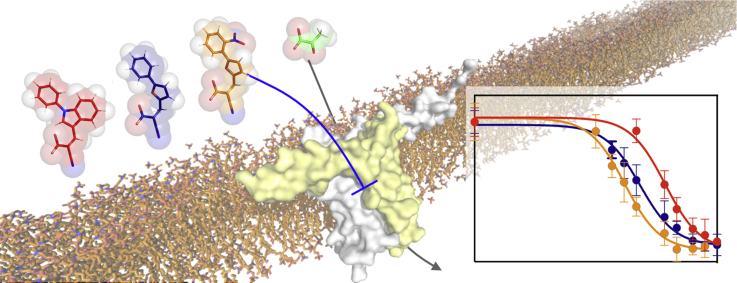
Objective: The mitochondrial pyruvate carrier (MPC) has emerged as a promising drug target for metabolic disorders, including non-alcoholic steatohepatitis and diabetes, metabolically dependent cancers and neurodegenerative diseases. A range of structurally diverse small molecule inhibitors have been proposed, but the nature of their interaction with MPC is not understood, and the composition of the functional human MPC is still debated. The goal of this study was to characterise the human MPC protein in vitro, to understand the chemical features that determine binding of structurally diverse inhibitors and to develop novel higher affinity ones.
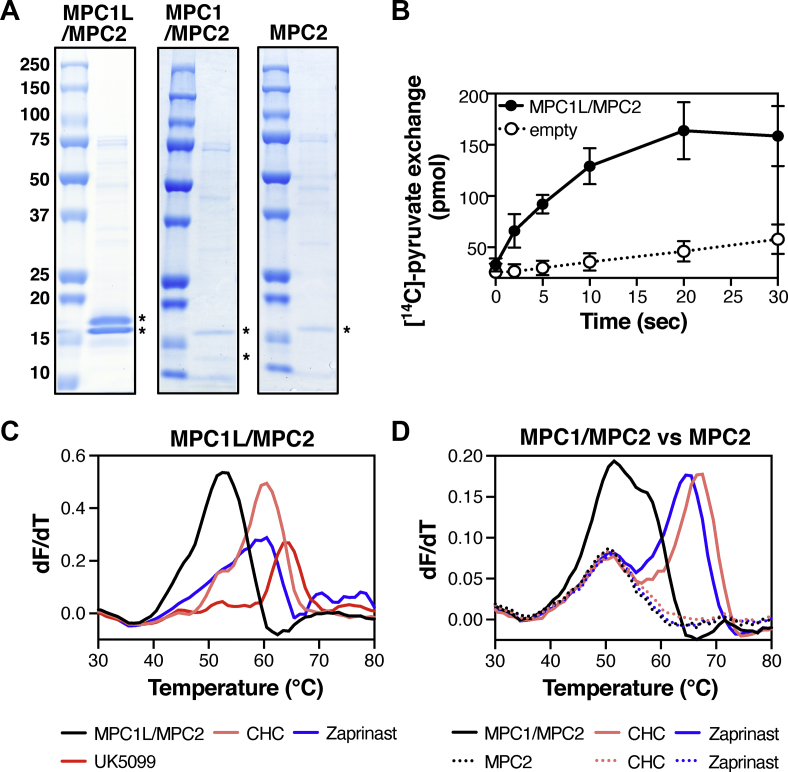
Methods: We recombinantly expressed and purified human MPC hetero-complexes and studied their composition, transport and inhibitor binding properties by establishing in vitro transport assays, high throughput thermostability shift assays and pharmacophore modeling.
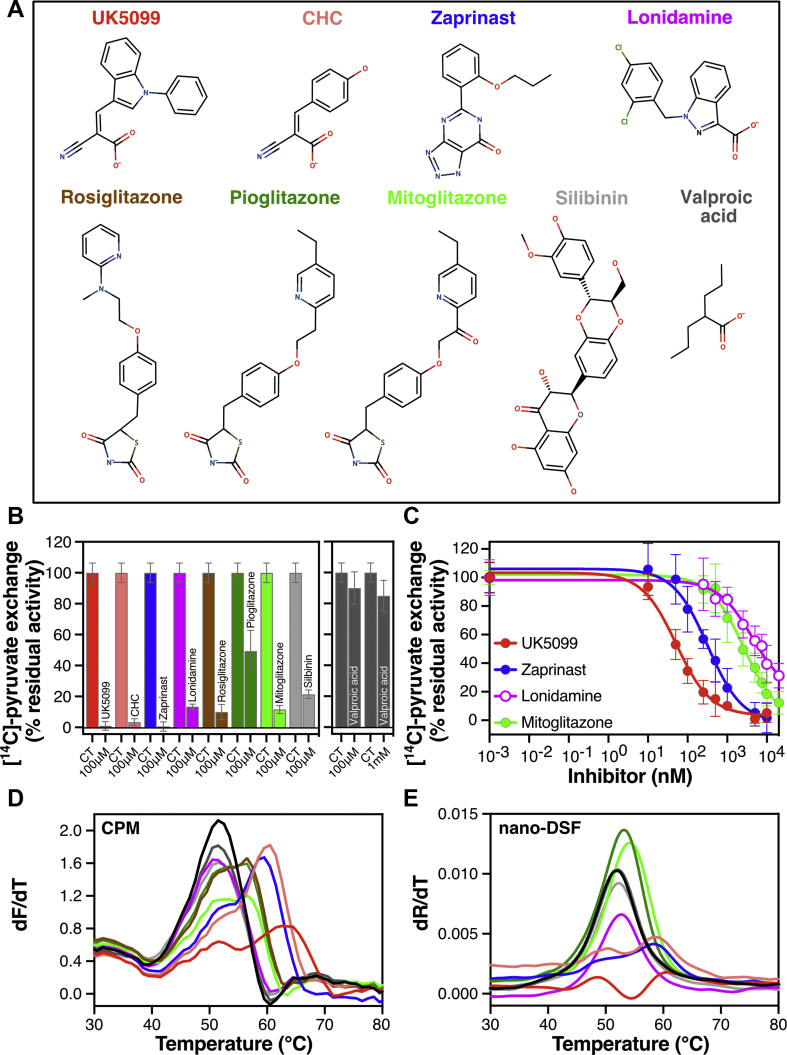
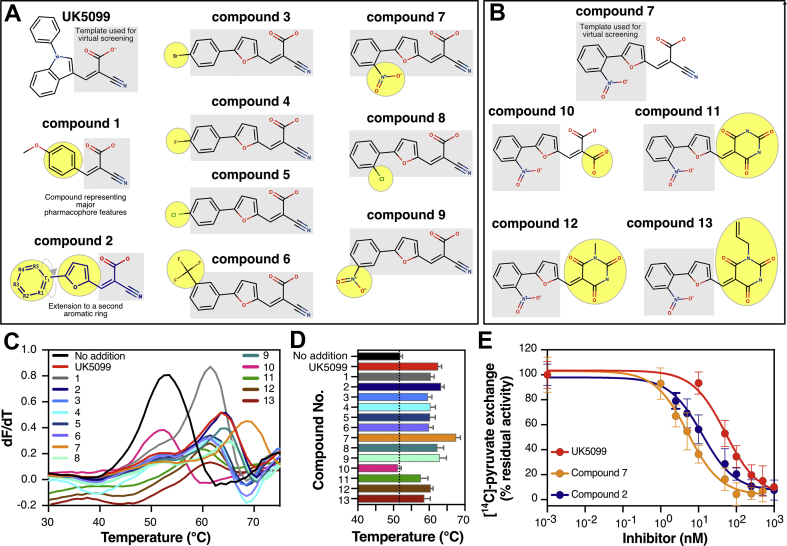
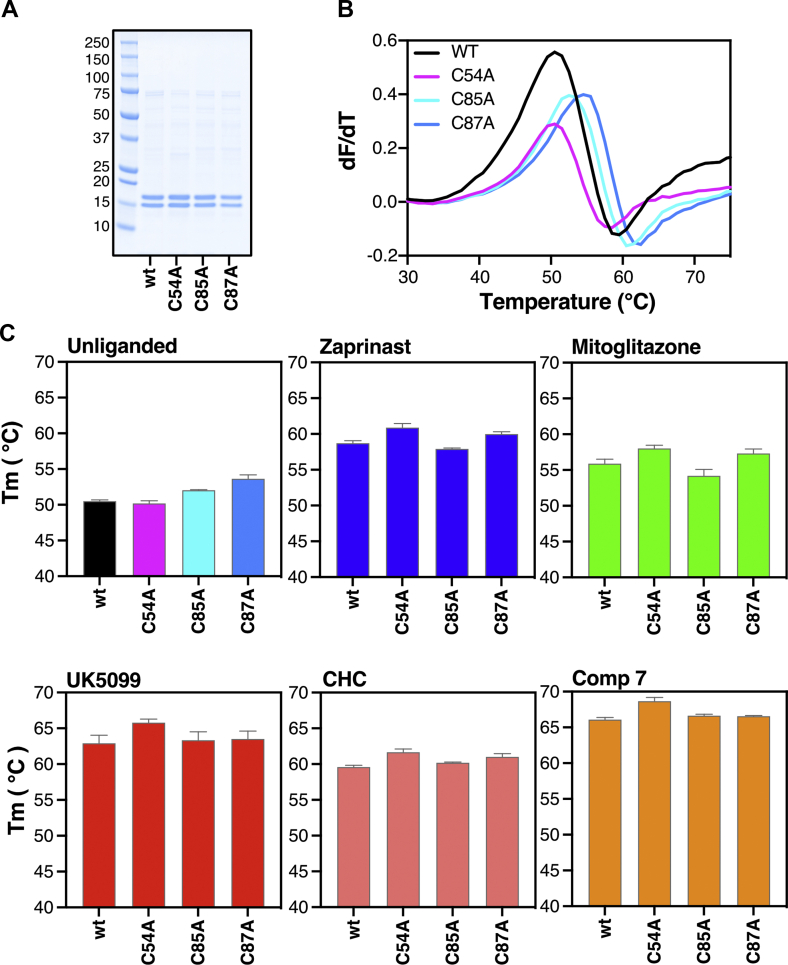
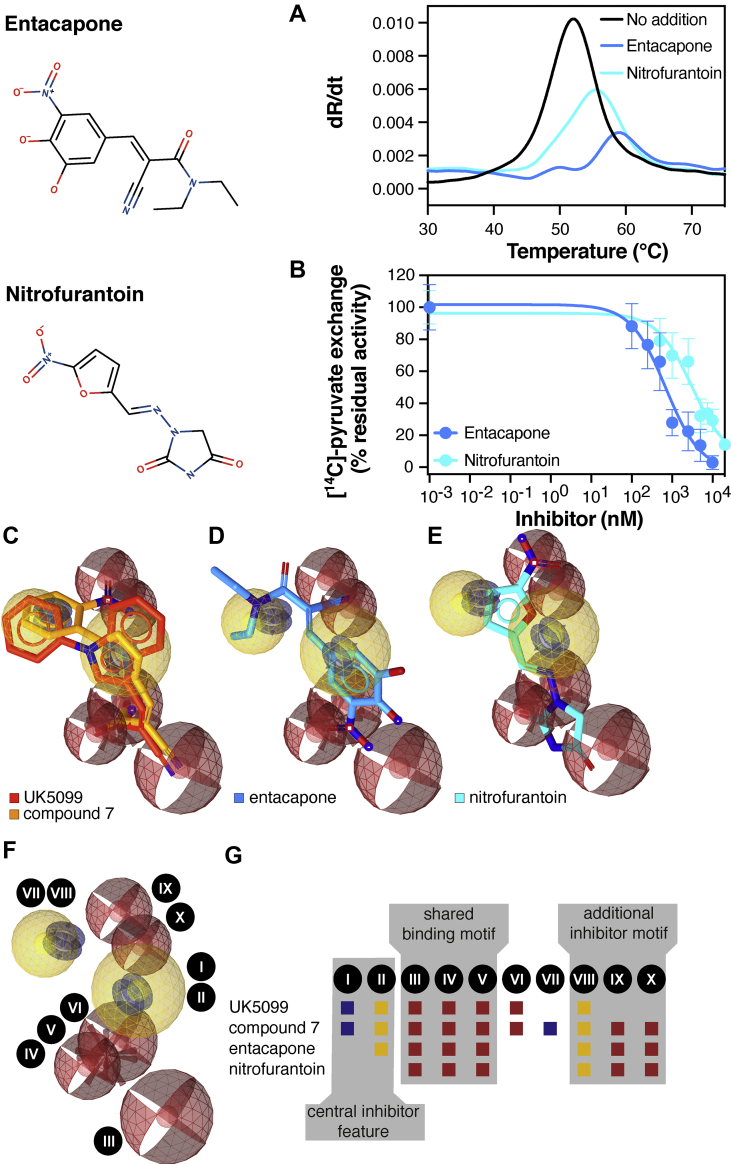
Results: We determined that the functional unit of human MPC is a hetero-dimer. We compared all different classes of MPC inhibitors to find that three closely arranged hydrogen bond acceptors followed by an aromatic ring are shared characteristics of all inhibitors and represent the minimal requirement for high potency. We also demonstrated that high affinity binding is not attributed to covalent bond formation with MPC cysteines, as previously proposed. Following the basic pharmacophore properties, we identified 14 new inhibitors of MPC, one outperforming compound UK5099 by tenfold. Two are the commonly prescribed drugs entacapone and nitrofurantoin, suggesting an off-target mechanism associated with their adverse effects.
Conclusions: This work defines the composition of human MPC and the essential MPC inhibitor characteristics. In combination with the functional assays we describe, this new understanding will accelerate the development of clinically relevant MPC modulators.
Keywords: Inhibition; Mitochondria; Mitochondrial pyruvate carrier; Mitochondrial transport; Small molecules.
Copyright © 2022 The Author(s). Published by Elsevier GmbH.. All rights reserved.
Figures
References
-
- Colca J.R., McDonald W.G., Adams W.J. MSDC-0602K, a metabolic modulator directed at the core pathology of non-alcoholic steatohepatitis. Expert Opinion on Investigational Drugs. 2018;27:631–636. - PubMed
-
- Harrison S.A., Alkhouri N., Davison B.A., Sanyal A., Edwards C., Colca J.R., et al. Insulin sensitizer MSDC-0602K in non-alcoholic steatohepatitis: a randomized, double-blind, placebo-controlled phase IIb study. Journal of Hepatology. 2020;72:613–626. - PubMed
-
- Ghosh A., Tyson T., George S., Hildebrandt E.N., Steiner J.A., Madaj Z., et al. Mitochondrial pyruvate carrier regulates autophagy, inflammation, and neurodegeneration in experimental models of Parkinson's disease. Science Translational Medicine. 2016;8 368ra174. - PubMed
Publication types
MeSH terms
Substances
Associated data
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
