

Mitochondrial Dynamics, Mitophagy, and Mitochondria-Endoplasmic Reticulum Contact Sites Crosstalk Under Hypoxia
- PMID: 35281107
- PMCID: PMC8914053
- DOI: 10.3389/fcell.2022.848214
Mitochondrial Dynamics, Mitophagy, and Mitochondria-Endoplasmic Reticulum Contact Sites Crosstalk Under Hypoxia
Abstract
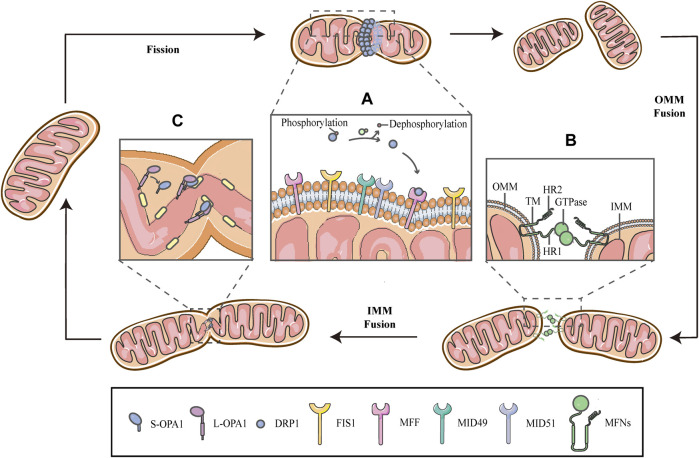
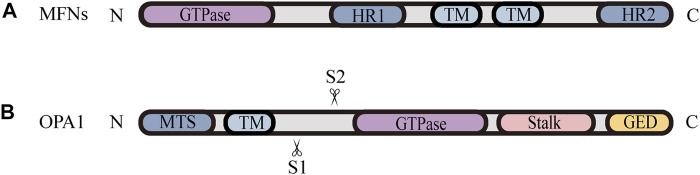
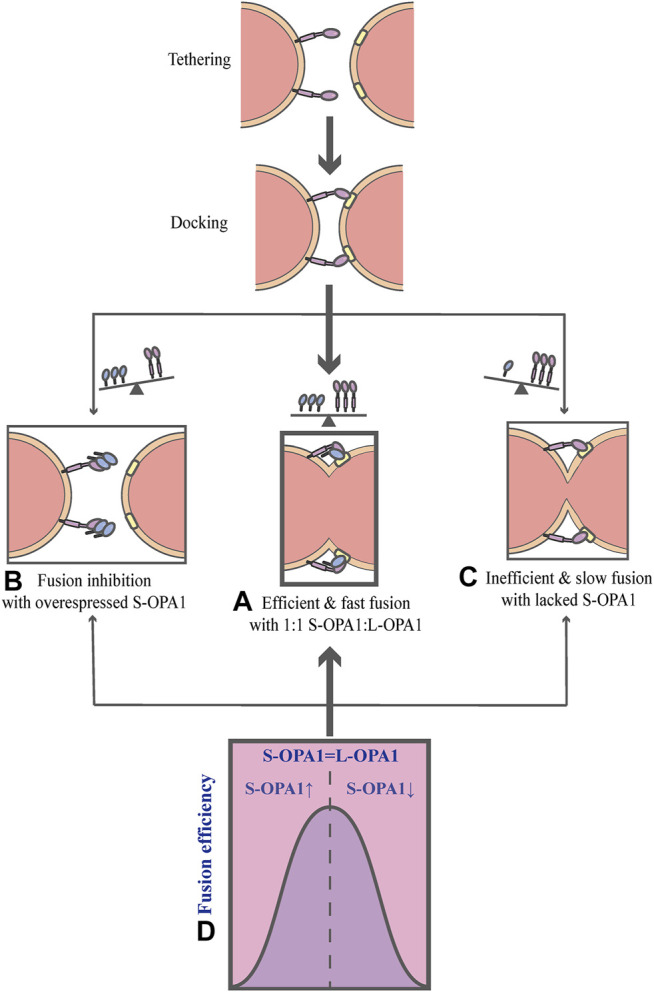
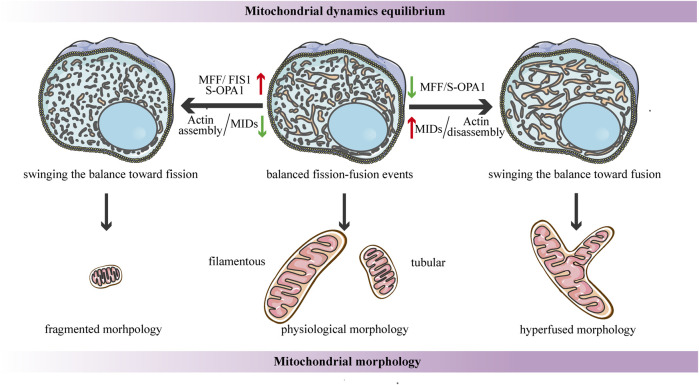
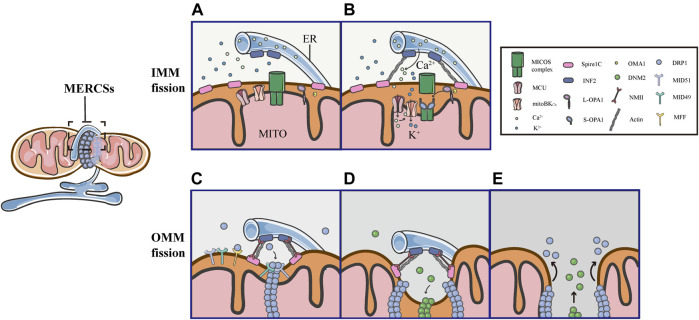
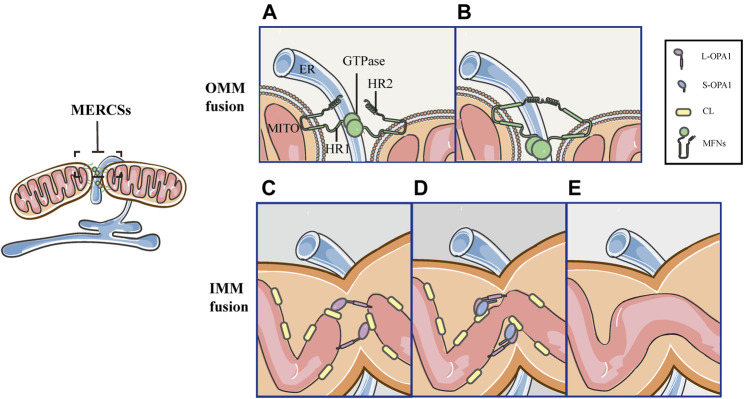
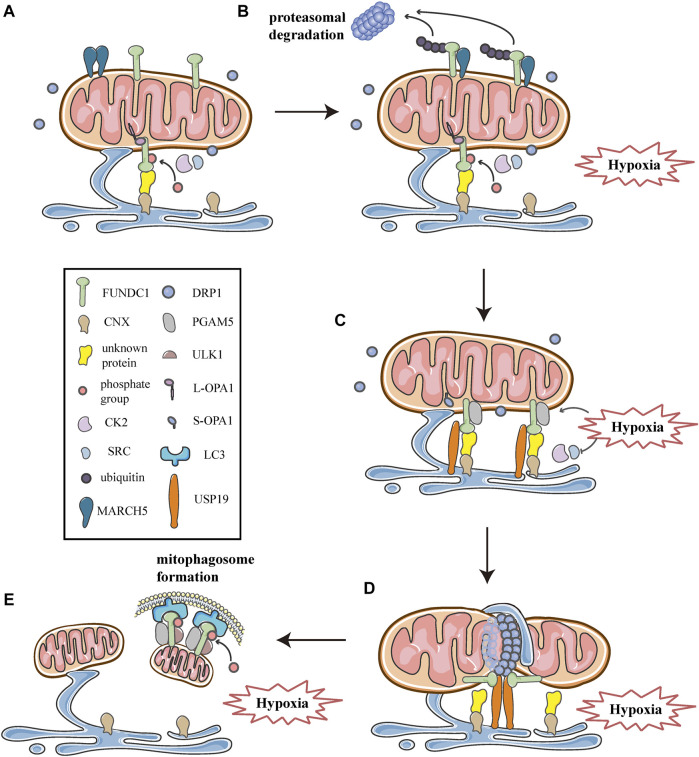
Mitochondria are double membrane organelles within eukaryotic cells, which act as cellular power houses, depending on the continuous availability of oxygen. Nevertheless, under hypoxia, metabolic disorders disturb the steady-state of mitochondrial network, which leads to dysfunction of mitochondria, producing a large amount of reactive oxygen species that cause further damage to cells. Compelling evidence suggests that the dysfunction of mitochondria under hypoxia is linked to a wide spectrum of human diseases, including obstructive sleep apnea, diabetes, cancer and cardiovascular disorders. The functional dichotomy of mitochondria instructs the necessity of a quality-control mechanism to ensure a requisite number of functional mitochondria that are present to fit cell needs. Mitochondrial dynamics plays a central role in monitoring the condition of mitochondrial quality. The fission-fusion cycle is regulated to attain a dynamic equilibrium under normal conditions, however, it is disrupted under hypoxia, resulting in mitochondrial fission and selective removal of impaired mitochondria by mitophagy. Current researches suggest that the molecular machinery underlying these well-orchestrated processes are coordinated at mitochondria-endoplasmic reticulum contact sites. Here, we establish a holistic understanding of how mitochondrial dynamics and mitophagy are regulated at mitochondria-endoplasmic reticulum contact sites under hypoxia.
Keywords: hypoxia; mitochondria; mitochondria-endoplasmic reticulum contact sites; mitochondrial dynamics; mitophagy.
Copyright © 2022 Wang, Tan, Miao and Zhang.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
-
- Baburamani A., Hurling C., Stolp H., Sobotka K., Gressens P., Hagberg H., et al. (2015). Chloe Hurling, Helen Stolp, Kristina Sobotka, Pierre Gressens, Henrik Hagberg, and Claire ThorntonMitochondrial Optic Atrophy (OPA) 1 Processing Is Altered in Response to Neonatal Hypoxic-Ischemic Brain Injury. Ijms 16 (9), 22509–22526. 10.3390/ijms160922509 - DOI - PMC - PubMed
Publication types
LinkOut - more resources
Full Text Sources