

Selective Recognition of Carbohydrate Antigens by Germline Antibodies Isolated from AID Knockout Mice
- PMID: 35282679
- PMCID: PMC10506689
- DOI: 10.1021/jacs.1c12745
Selective Recognition of Carbohydrate Antigens by Germline Antibodies Isolated from AID Knockout Mice
Abstract
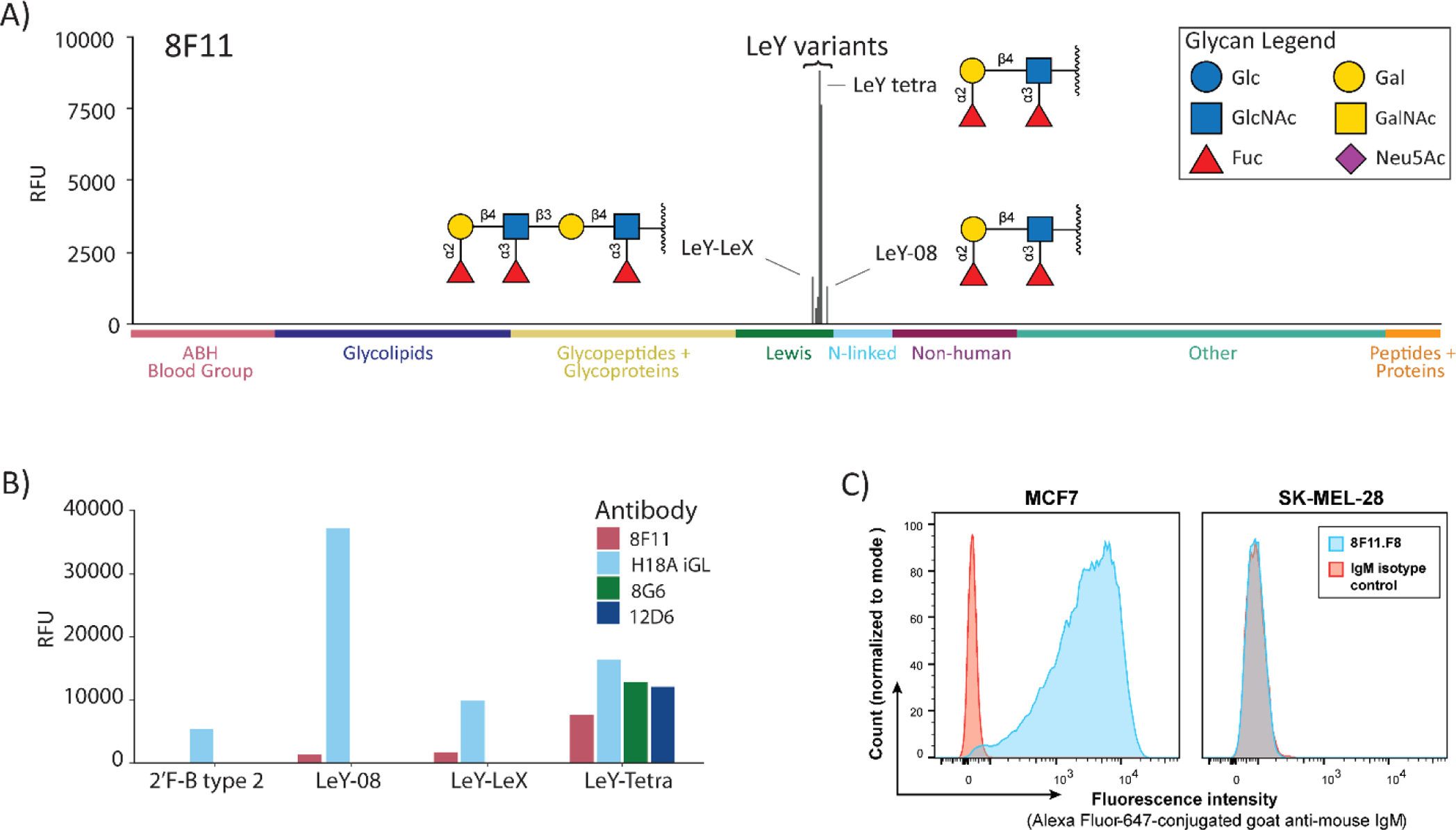
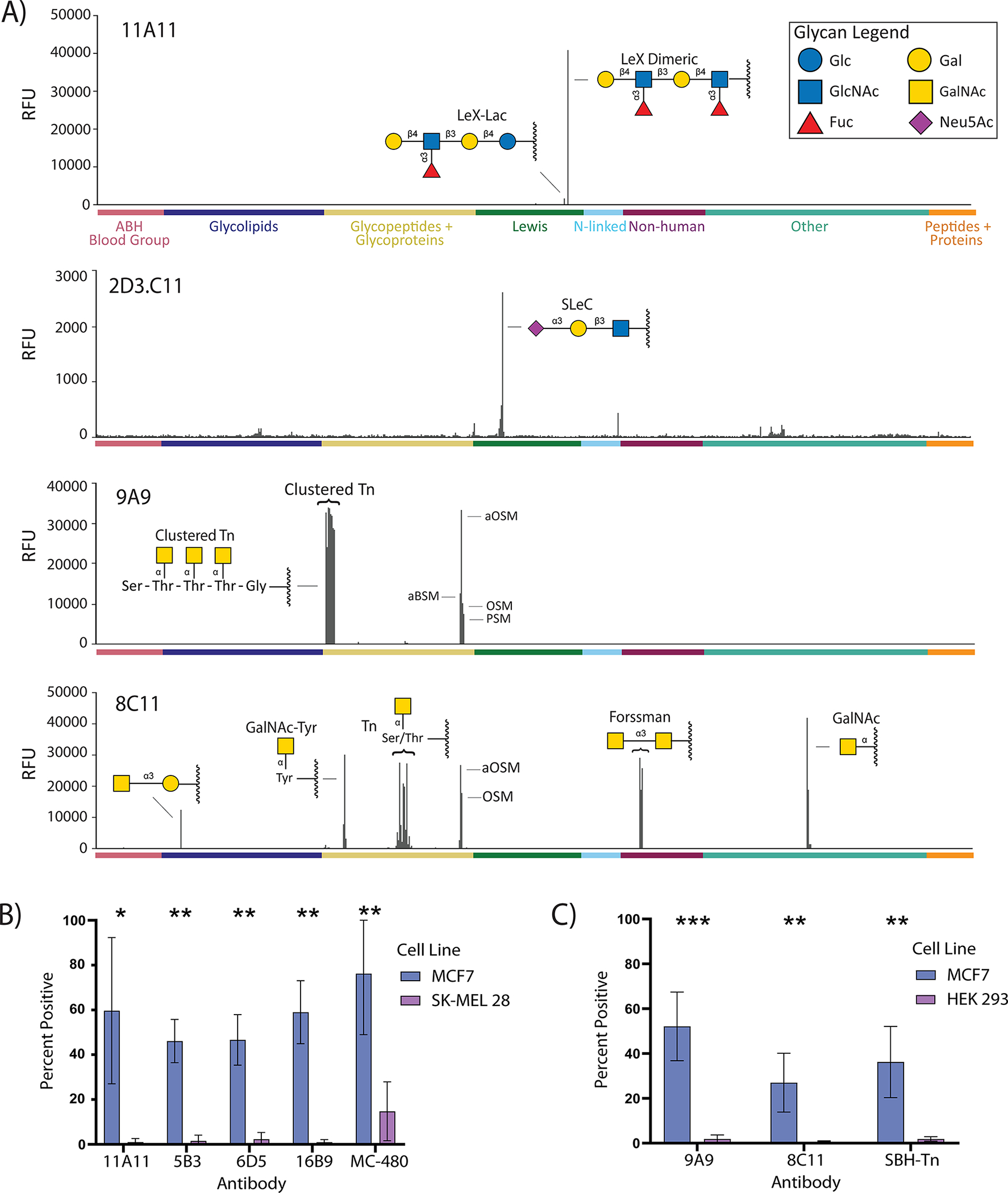
Germline antibodies, the initial set of antibodies produced by the immune system, are critical for host defense, and information about their binding properties can be useful for designing vaccines, understanding the origins of autoantibodies, and developing monoclonal antibodies. Numerous studies have found that germline antibodies are polyreactive with malleable, flexible binding pockets. While insightful, it remains unclear how broadly this model applies, as there are many families of antibodies that have not yet been studied. In addition, the methods used to obtain germline antibodies typically rely on assumptions and do not work well for many antibodies. Herein, we present a distinct approach for isolating germline antibodies that involves immunizing activation-induced cytidine deaminase (AID) knockout mice. This strategy amplifies antigen-specific B cells, but somatic hypermutation does not occur because AID is absent. Using synthetic haptens, glycoproteins, and whole cells, we obtained germline antibodies to an assortment of clinically important tumor-associated carbohydrate antigens, including Lewis Y, the Tn antigen, sialyl Lewis C, and Lewis X (CD15/SSEA-1). Through glycan microarray profiling and cell binding, we demonstrate that all but one of these germline antibodies had high selectivity for their glycan targets. Using molecular dynamics simulations, we provide insights into the structural basis of glycan recognition. The results have important implications for designing carbohydrate-based vaccines, developing anti-glycan monoclonal antibodies, and understanding antibody evolution within the immune system.
Conflict of interest statement
The authors declare no competing financial interest.
Figures







References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
