

The Unfolded Protein Responses in Health, Aging, and Neurodegeneration: Recent Advances and Future Considerations
- PMID: 35283733
- PMCID: PMC8914544
- DOI: 10.3389/fnmol.2022.831116
The Unfolded Protein Responses in Health, Aging, and Neurodegeneration: Recent Advances and Future Considerations
Abstract
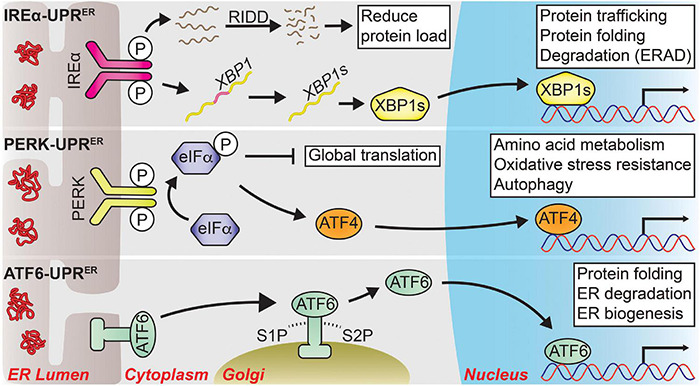
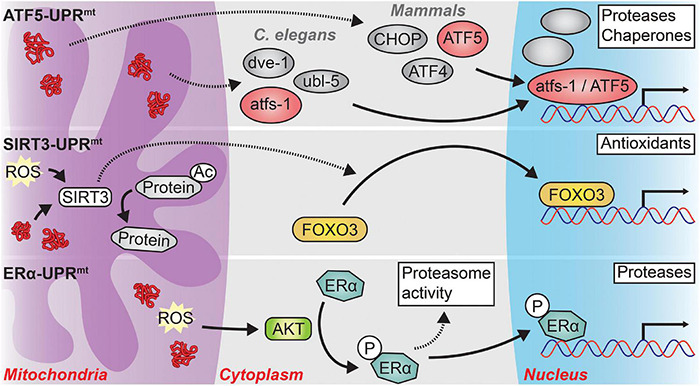
Aging and age-related neurodegeneration are both associated with the accumulation of unfolded and abnormally folded proteins, highlighting the importance of protein homeostasis (termed proteostasis) in maintaining organismal health. To this end, two cellular compartments with essential protein folding functions, the endoplasmic reticulum (ER) and the mitochondria, are equipped with unique protein stress responses, known as the ER unfolded protein response (UPR ER ) and the mitochondrial UPR (UPR mt ), respectively. These organellar UPRs play roles in shaping the cellular responses to proteostatic stress that occurs in aging and age-related neurodegeneration. The loss of adaptive UPR ER and UPR mt signaling potency with age contributes to a feed-forward cycle of increasing protein stress and cellular dysfunction. Likewise, UPR ER and UPR mt signaling is often altered in age-related neurodegenerative diseases; however, whether these changes counteract or contribute to the disease pathology appears to be context dependent. Intriguingly, altering organellar UPR signaling in animal models can reduce the pathological consequences of aging and neurodegeneration which has prompted clinical investigations of UPR signaling modulators as therapeutics. Here, we review the physiology of both the UPR ER and the UPR mt , discuss how UPR ER and UPR mt signaling changes in the context of aging and neurodegeneration, and highlight therapeutic strategies targeting the UPR ER and UPR mt that may improve human health.
Keywords: aging; endoplasmic reticulum unfolded protein response; mitochondrial unfolded protein response (UPRmt); neurodegeneration; unfolded protein response (UPR).
Copyright © 2022 Wodrich, Scott, Shukla, Harris and Giniger.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures
References
Publication types
LinkOut - more resources
Full Text Sources