

Multi-Approach Analysis Reveals Pathways of Cold Tolerance Divergence in Camellia japonica
- PMID: 35283896
- PMCID: PMC8914472
- DOI: 10.3389/fpls.2022.811791
Multi-Approach Analysis Reveals Pathways of Cold Tolerance Divergence in Camellia japonica
Abstract
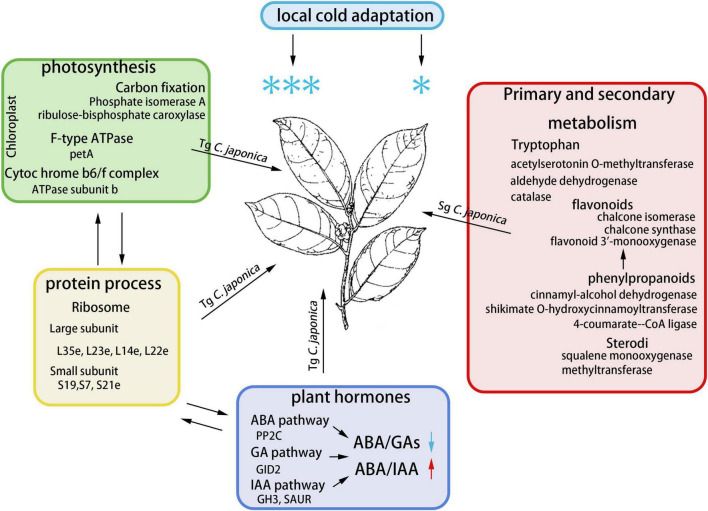
Understanding the molecular mechanism of the cold response is critical to improve horticultural plant cold tolerance. Here, we documented the physiological, transcriptome, proteome, and hormonal dynamics to cold stress in temperate genotype (Tg) and subtropical genotype (Sg) populations of Camellia japonica. Tg C. japonica suffered minimal osmotic and oxidative damage compared to Sg C. japonica under the same cold treatment. Transcriptional and translational differences increased under the cold treatment, indicating that Tg C. japonica was affected by the environment and displayed both conserved and divergent mechanisms. About 60% of the genes responding to cold had similar dynamics in the two populations, but 1,896 transcripts and 455 proteins differentially accumulated in response to the cold between Tg and Sg C. japonica. Co-expression analysis showed that the ribosomal protein and genes related to photosynthesis were upregulated in Tg C. japonica, and tryptophan, phenylpropanoid, and flavonoid metabolism were regulated differently between the two populations under cold stress. The divergence of these genes reflected a difference in cold responsiveness. In addition, the decrease in the abscisic acid (ABA)/gibberellic acid (GA) ratio regulated by biosynthetic signal transduction pathway enhanced cold resistance in Tg C. japonica, suggesting that hormones may regulate the difference in cold responsiveness. These results provide a new understanding of the molecular mechanism of cold stress and will improve cold tolerance in horticultural plants.
Keywords: Camellia japonica; co-expression; cold; plant hormone; proteome; transcriptome.
Copyright © 2022 Fan, Zhang, Li, Wu, Yang, Yin, Liu, Fan and Li.
Conflict of interest statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Figures






References
-
- Batista-Santos P., Lidon F. C., Fortunato A., Leitão A. E., Lopes E., Partelli F., et al. (2011). The impact of cold on photosynthesis in genotypes of Coffea spp.-Photosystem sensitivity, photoprotective mechanisms and gene expression. J. Plant Physiol. 168 792–806. 10.1016/j.jplph.2010.11.013 - DOI - PubMed
-
- Charles S. A., Halliwell B. (1981). The role of calcium ions and the thioredoxin system in regulation of spinach chloroplast fructosebisphosphatase. Cell Calcium. 2 211–224. 10.1016/0143-4160(81)90037-3 - DOI
LinkOut - more resources
Full Text Sources
Miscellaneous