

Mechanistic contributions of Kupffer cells and liver sinusoidal endothelial cells in nanoparticle-induced antigen-specific immune tolerance
- PMID: 35286851
- PMCID: PMC11225973
- DOI: 10.1016/j.biomaterials.2022.121457
Mechanistic contributions of Kupffer cells and liver sinusoidal endothelial cells in nanoparticle-induced antigen-specific immune tolerance
Abstract
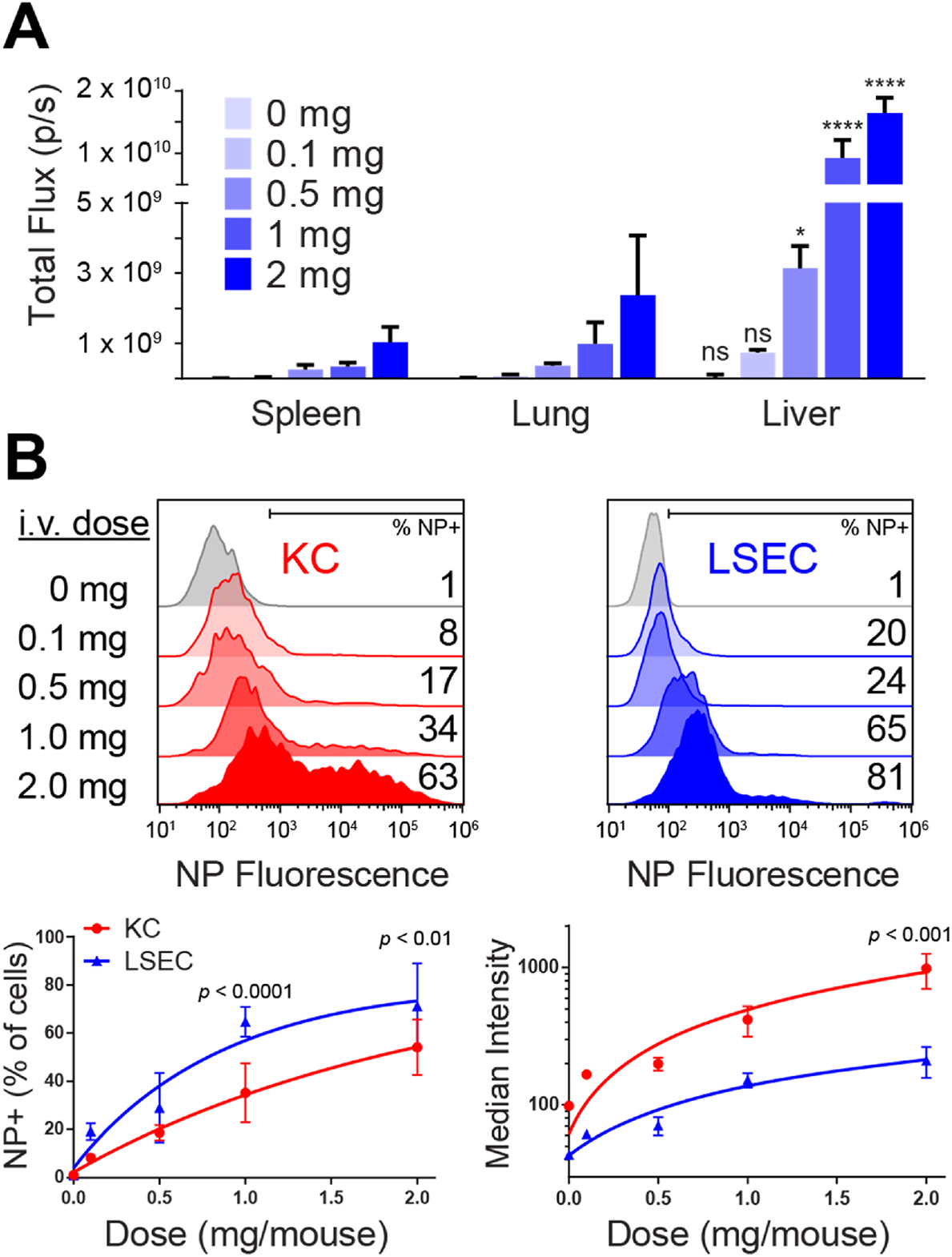
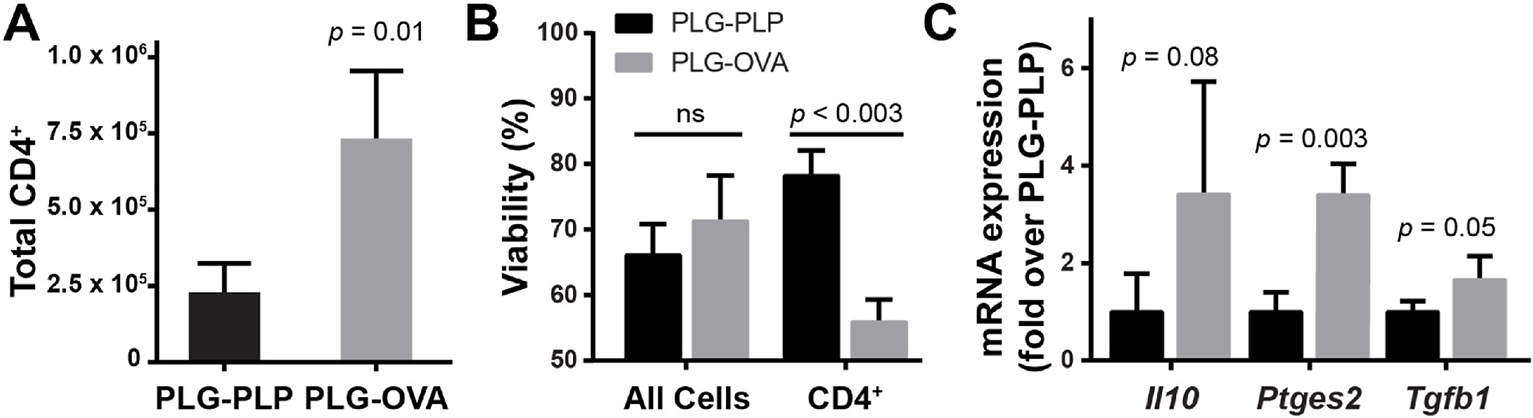
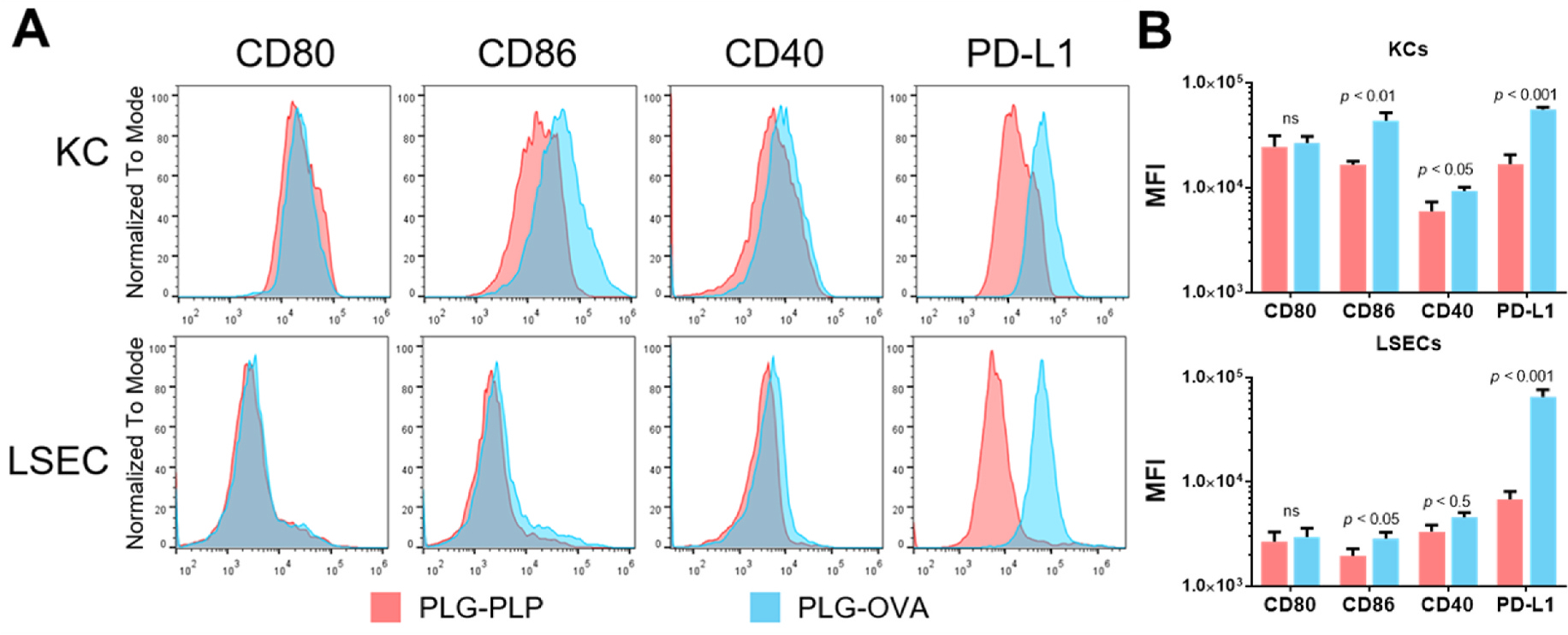
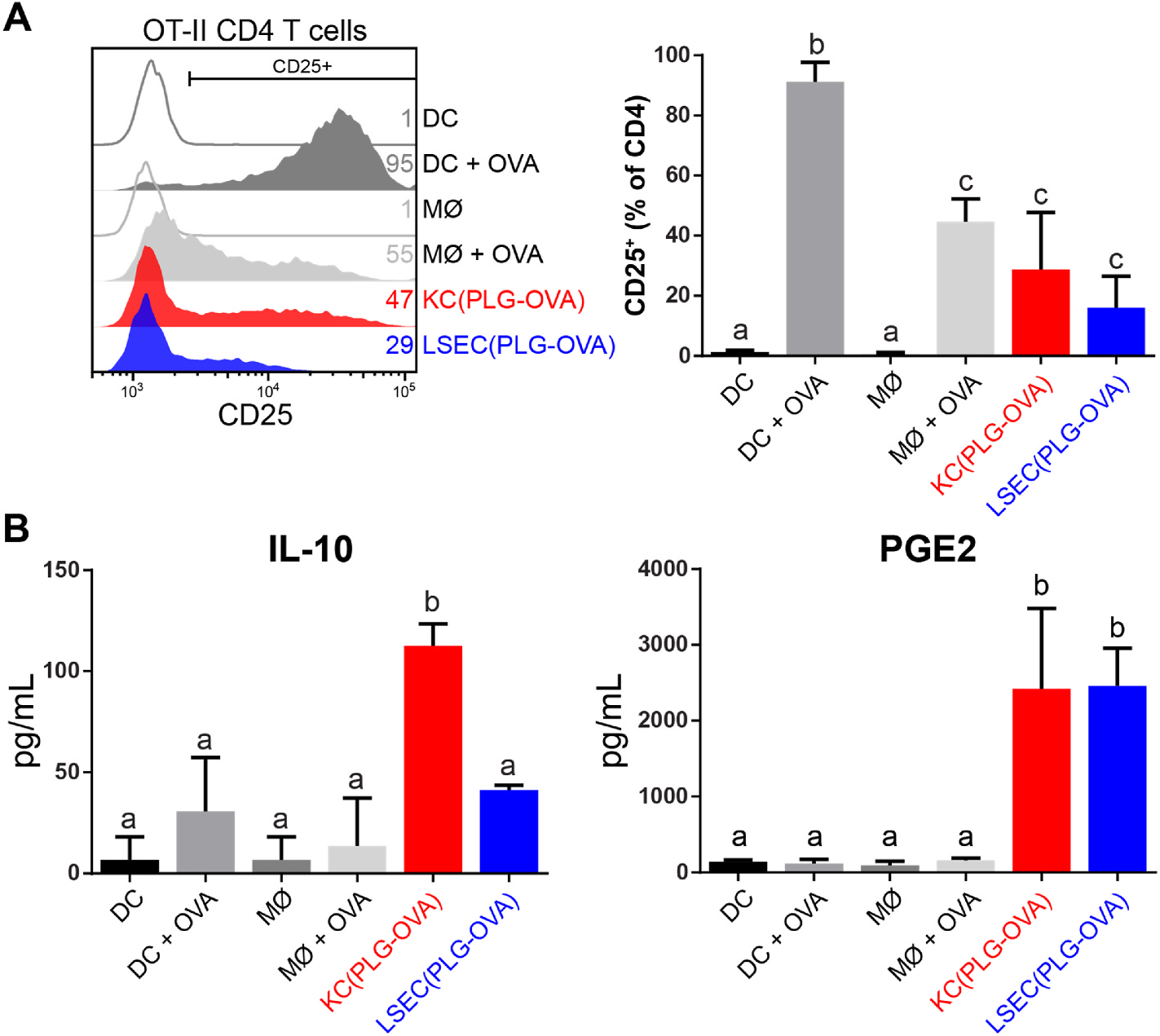
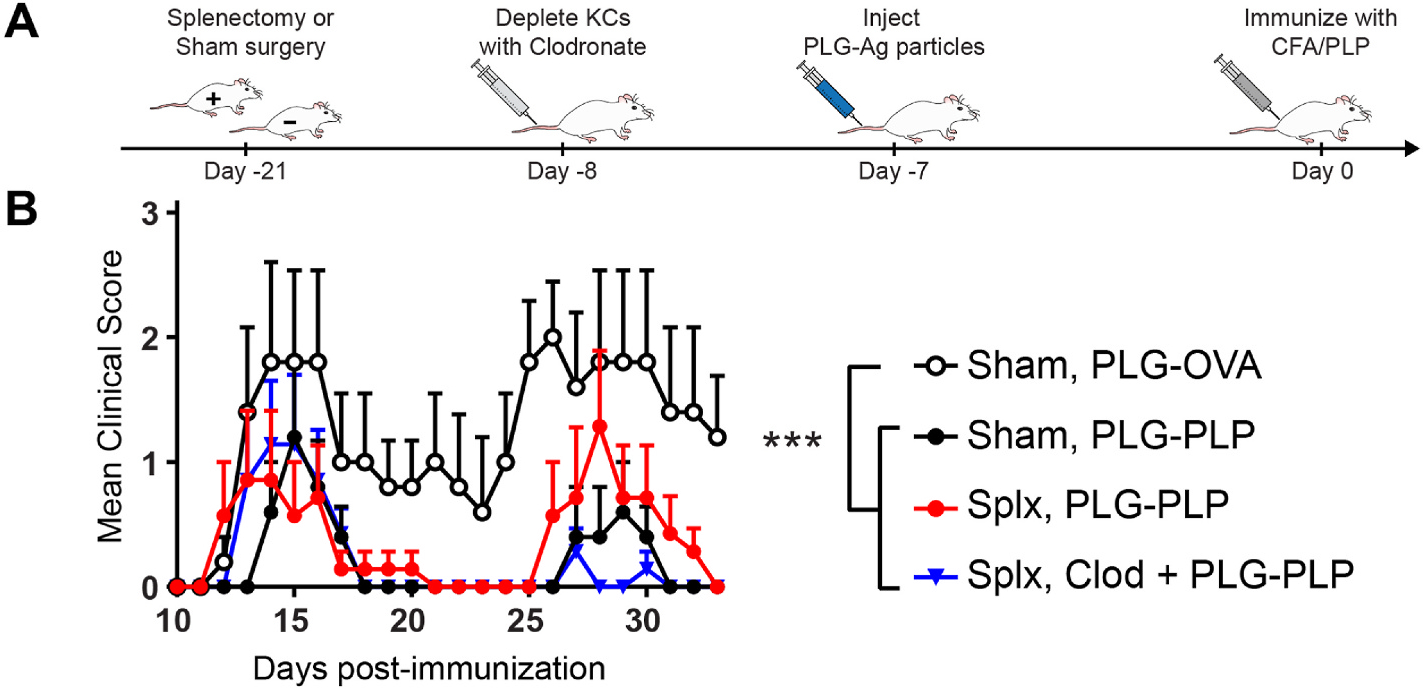
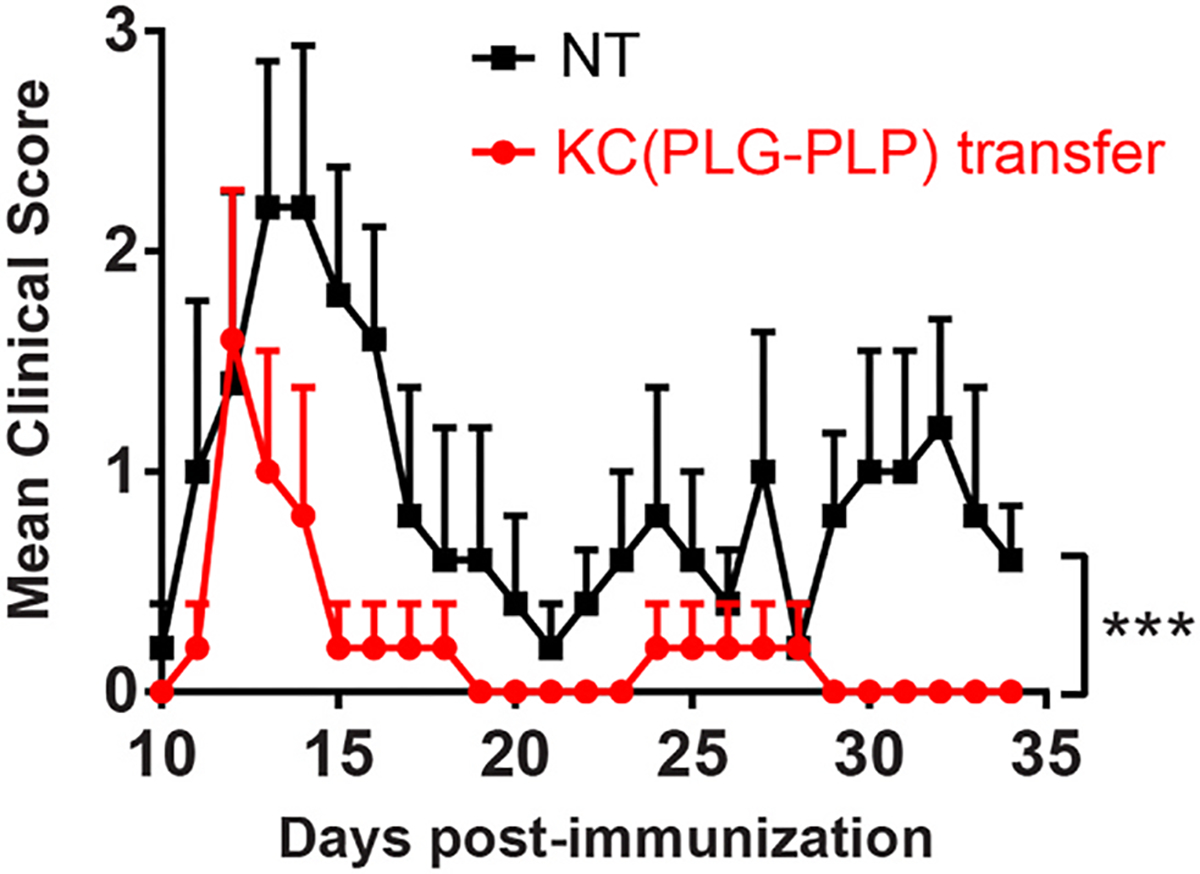
The intravenous delivery of disease-relevant antigens (Ag) by polymeric nanoparticles (NP-Ags) has demonstrated Ag-specific immune tolerance in autoimmune and allergic disorders as well as allogeneic transplant rejection. NP-Ags are observed to distribute to the spleen, which has an established role in the induction of immune tolerance. However, studies have shown that the spleen is dispensable for NP-Ag-induced tolerance, suggesting significant contributions from other immunological sites. Here, we investigated the tolerogenic contributions of Kupffer cells (KCs) and liver sinusoidal endothelial cells (LSECs) to NP-Ag-induced tolerance in a mouse model of multiple sclerosis, experimental autoimmune encephalomyelitis (EAE). Intravenously delivered Ag-conjugated poly(lactide-co-glycolide) NPs (PLG-Ag) distributed largely to the liver, where they associated with both KCs and LSECs. This distribution was accompanied by CD4 T cell accumulation, clonal deletion, and PD-L1 expression by KCs and LSECs. Ex vivo co-cultures of PLG-Ag-treated KCs or LSECs with Ag-specific CD4 T cells resulted in PGE2 and IL-10 or PGE2 secretion, respectively. KC depletion and adoptive transfer experiments demonstrated that KCs were sufficient, but not necessary, to mediate PLG-Ag-induced tolerance in EAE. The durability of PLG-Ag-induced tolerance in the absence of KCs may be attributed to the distribution of PLG-Ags to LSECs, which demonstrated similar levels of PD-L1, PGE2, and T cell stimulatory ability. Collectively, these studies provide mechanistic support for the role of liver KCs and LSECs in Ag-specific tolerance for a biomaterial platform that is currently being evaluated in clinical trials.
Keywords: Immune tolerance; Immunomodulation; Kupffer cells; Liver sinusoidal endothelial cells; Nanoparticles; PLG.
Copyright © 2022 Elsevier Ltd. All rights reserved.
Conflict of interest statement
Declaration of competing interest
The authors declare the following financial interests/personal relationships which may be considered as potential competing interests:
Lonnie Shea reports a relationship with Cour Pharmaceuticals Development Company that includes: consulting or advisory and funding grants. Ryan Pearson reports a relationship with Cour Pharmaceuticals Development Company that includes: consulting or advisory. Stephen Miller reports a relationship with Cour Pharmaceuticals Development Company that includes: consulting or advisory and funding grants.
Figures
Similar articles
-
An antigen-encapsulating nanoparticle platform for TH1/17 immune tolerance therapy.Nanomedicine. 2017 Jan;13(1):191-200. doi: 10.1016/j.nano.2016.09.007. Epub 2016 Oct 6. Nanomedicine. 2017. PMID: 27720992 Free PMC article.
-
Nanoparticle dose and antigen loading attenuate antigen-specific T-cell responses.Biotechnol Bioeng. 2023 Jan;120(1):284-296. doi: 10.1002/bit.28252. Epub 2022 Oct 21. Biotechnol Bioeng. 2023. PMID: 36221192 Free PMC article.
-
Enhancement of the Tolerogenic Phenotype in the Liver by ImmTOR Nanoparticles.Front Immunol. 2021 May 25;12:637469. doi: 10.3389/fimmu.2021.637469. eCollection 2021. Front Immunol. 2021. PMID: 34113339 Free PMC article.
-
Immune tolerance: what is unique about the liver.J Autoimmun. 2010 Feb;34(1):1-6. doi: 10.1016/j.jaut.2009.08.008. Epub 2009 Aug 29. J Autoimmun. 2010. PMID: 19717280 Review.
-
Role of Liver-Mediated Tolerance in Nanoparticle-Based Tumor Therapy.Cells. 2020 Aug 28;9(9):1985. doi: 10.3390/cells9091985. Cells. 2020. PMID: 32872352 Free PMC article. Review.
Cited by
-
Development of protein-polymer conjugate nanoparticles for modulation of dendritic cell phenotype and antigen-specific CD4 T cell responses.ACS Appl Polym Mater. 2023 Nov 10;5(11):8794-8807. doi: 10.1021/acsapm.3c00548. Epub 2023 Oct 9. ACS Appl Polym Mater. 2023. PMID: 38911349 Free PMC article.
-
Vaccines for immune tolerance against autoimmune disease.Adv Drug Deliv Rev. 2023 Dec;203:115140. doi: 10.1016/j.addr.2023.115140. Epub 2023 Nov 18. Adv Drug Deliv Rev. 2023. PMID: 37980949 Free PMC article. Review.
-
Nanomaterials for antigen-specific immune tolerance therapy.Drug Deliv Transl Res. 2023 Jul;13(7):1859-1881. doi: 10.1007/s13346-022-01233-3. Epub 2022 Sep 12. Drug Deliv Transl Res. 2023. PMID: 36094655 Review.
-
Role of liver sinusoidal endothelial cell in metabolic dysfunction-associated fatty liver disease.Cell Commun Signal. 2024 Jun 28;22(1):346. doi: 10.1186/s12964-024-01720-9. Cell Commun Signal. 2024. PMID: 38943171 Free PMC article. Review.
-
Lipid-Polymer Hybrid Nanoparticles Utilize B Cells and Dendritic Cells to Elicit Distinct Antigen-Specific CD4+ and CD8+ T Cell Responses.ACS Appl Bio Mater. 2024 Aug 19;7(8):4818-4830. doi: 10.1021/acsabm.3c00229. Epub 2023 May 23. ACS Appl Bio Mater. 2024. PMID: 37219857 Free PMC article. Review.
References
-
- Bach J-F, The effect of infections on susceptibility to autoimmune and allergic diseases, N. Engl. J. Med. 347 (2002) 911–920. - PubMed
-
- Galvão VR, Castells MC, Hypersensitivity to biological agents—updated diagnosis, management, and treatment, J. Allergy Clin. Immunol. Pract. 3 (2015) 175–185. - PubMed
-
- Calabrese L, Molloy E, Progressive multifocal leucoencephalopathy in the rheumatic diseases: assessing the risks of biological immunosuppressive therapies, Ann. Rheum. Dis. 67 (2008) iii64–iii65. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
