

Endothelial Cell Fate Determination: A Top Notch Job in Vascular Decision-Making
- PMID: 35288401
- PMCID: PMC9619357
- DOI: 10.1101/cshperspect.a041183
Endothelial Cell Fate Determination: A Top Notch Job in Vascular Decision-Making
Abstract
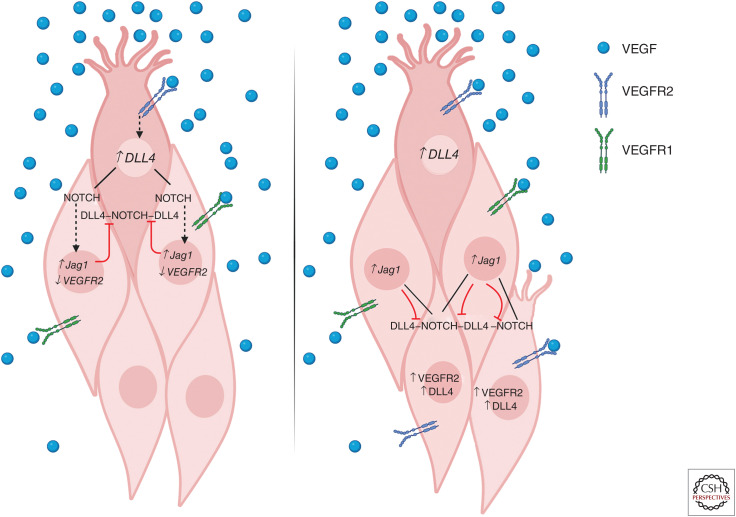
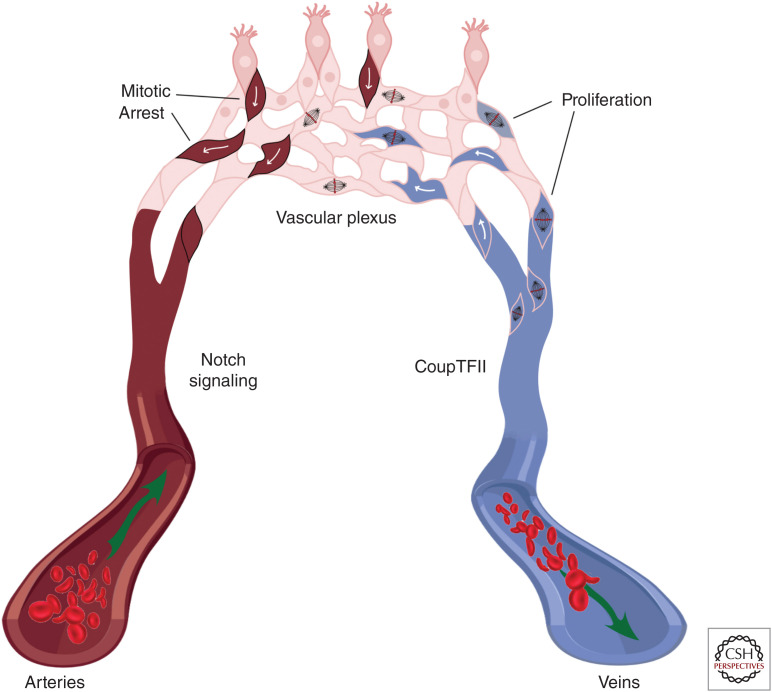
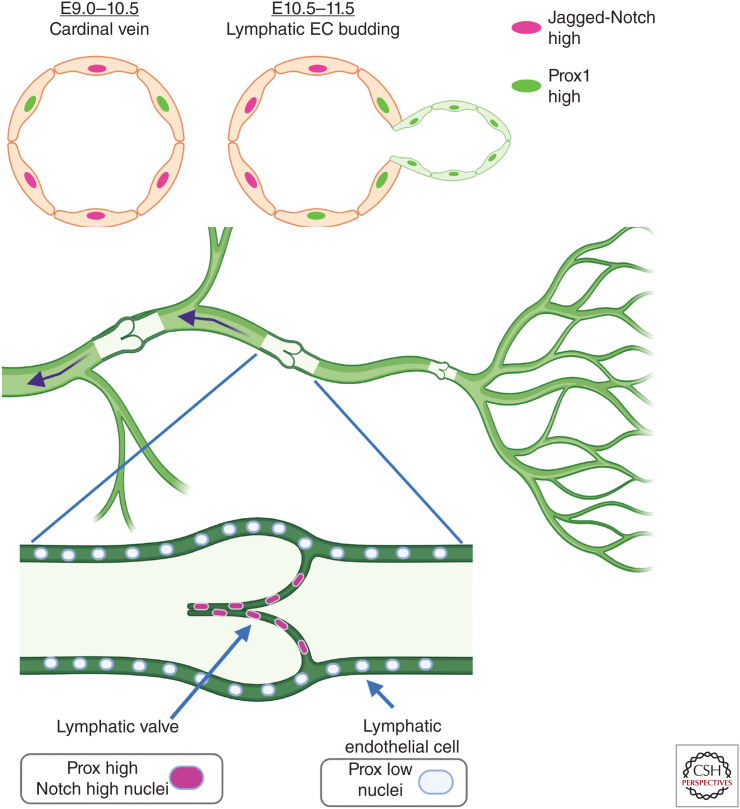
As vascular networks form, endothelial cells (ECs) undergo cell fate decisions that determine whether they become tip or stalk cells of the developing vascular plexus or mature into arterial, venous, or lymphatic endothelium. EC fate decisions are coordinated with neighboring cells to initiate sprouting, maintain endothelial barrier, or ensure appropriate specialization of vessels. We describe mechanisms that control EC fate at specific steps in these processes, with an emphasis on the role of the Notch signaling pathway. Specific EC fate determination steps that are highlighted are tip/stalk selection during sprouting angiogenesis, venous-arterial specification, arteriogenesis, lymphatic vessel specification, and lymphatic valve formation.
Copyright © 2022 Cold Spring Harbor Laboratory Press; all rights reserved.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials