

Oxidative metabolisms catalyzed Earth's oxygenation
- PMID: 35288554
- PMCID: PMC8921266
- DOI: 10.1038/s41467-022-28996-0
Oxidative metabolisms catalyzed Earth's oxygenation
Abstract
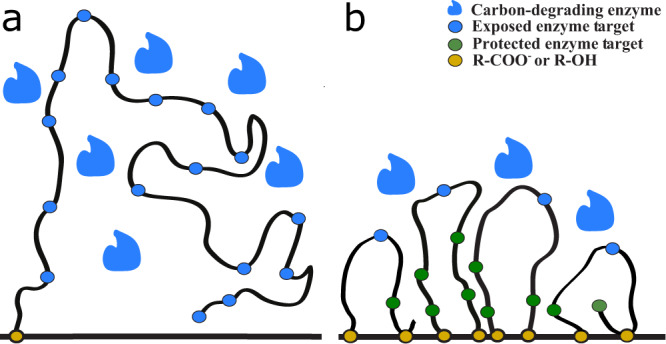
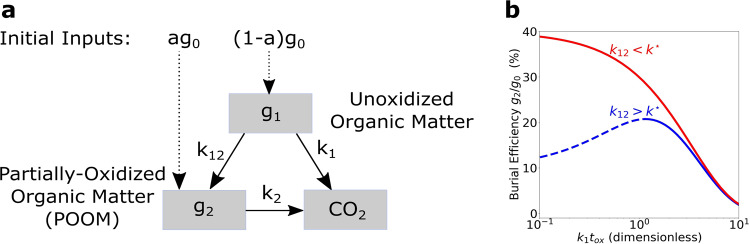
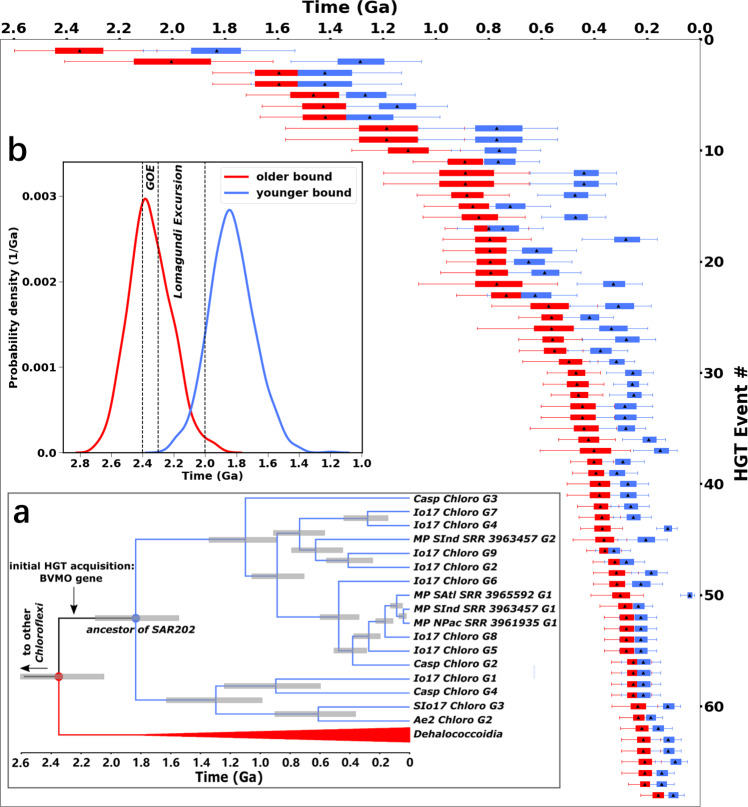
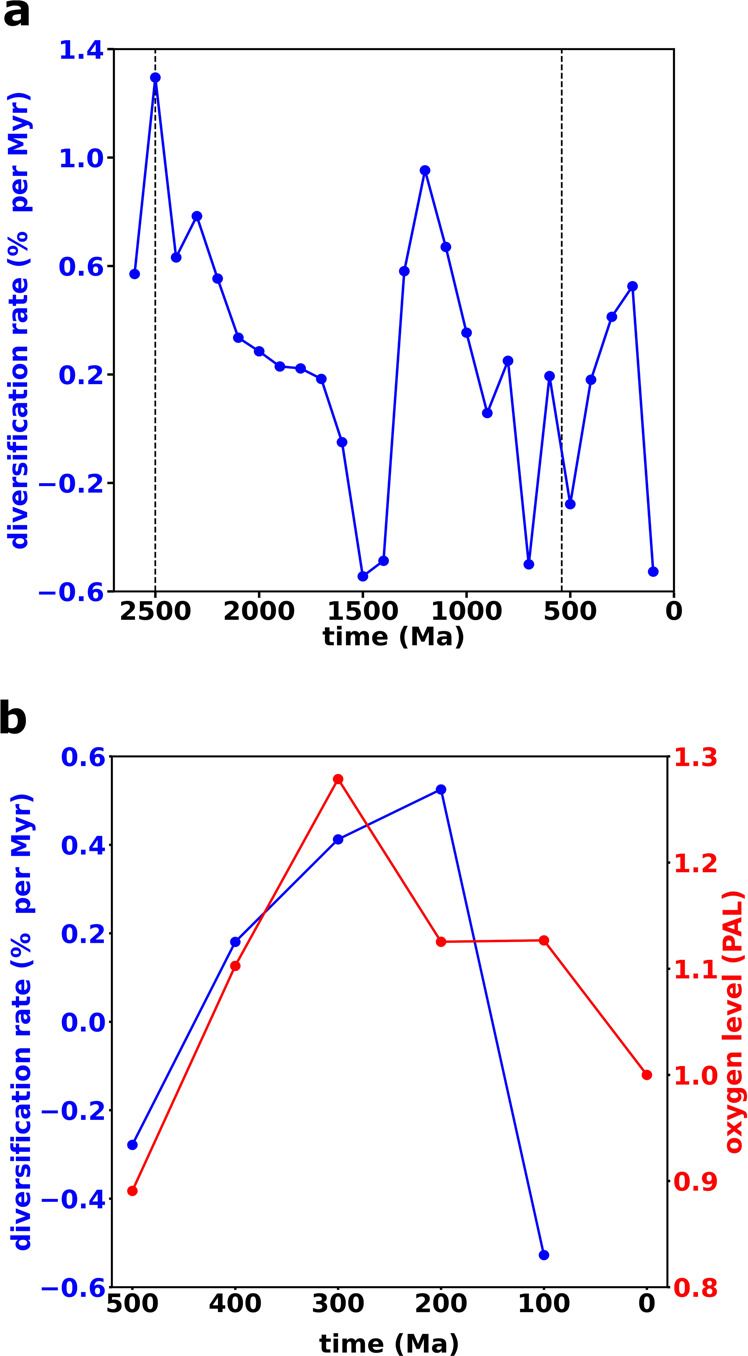
The burial of organic carbon, which prevents its remineralization via oxygen-consuming processes, is considered one of the causes of Earth's oxygenation. Yet, higher levels of oxygen are thought to inhibit burial. Here we propose a resolution of this conundrum, wherein Earth's initial oxygenation is favored by oxidative metabolisms generating partially oxidized organic matter (POOM), increasing burial via interaction with minerals in sediments. First, we introduce the POOM hypothesis via a mathematical argument. Second, we reconstruct the evolutionary history of one key enzyme family, flavin-dependent Baeyer-Villiger monooxygenases, that generates POOM, and show the temporal consistency of its diversification with the Proterozoic and Phanerozoic atmospheric oxygenation. Finally, we propose that the expansion of oxidative metabolisms instigated a positive feedback, which was amplified by the chemical changes to minerals on Earth's surface. Collectively, these results suggest that Earth's oxygenation is an autocatalytic transition induced by a combination of biological innovations and geological changes.
© 2022. The Author(s).
Conflict of interest statement
The authors declare no competing interests.
Figures
References
-
- Canfield, D. E. Oxygen: A Four Billion Year History (Princeton University Press, 2014).
-
- Lyons TW, Reinhard CT, Planavsky NJ. The rise of oxygen in Earth’s early ocean and atmosphere. Nature. 2014;506:307–315. - PubMed
-
- Catling DC, Claire MW. How Earth’s atmosphere evolved to an oxic state: a status report. Earth Planet. Sci. Lett. 2005;237:1–20.
-
- Catling DC, Zahnle KJ, McKay CP. Biogenic methane, hydrogen escape, and the irreversible oxidation of early Earth. Science. 2001;293:839–843. - PubMed
-
- Kump LR, Barley ME. Increased subaerial volcanism and the rise of atmospheric oxygen 2.5 billion years ago. Nature. 2007;448:1033–1036. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
