

Copper Induces Protein Aggregation, a Toxic Process Compensated by Molecular Chaperones
- PMID: 35289645
- PMCID: PMC9040851
- DOI: 10.1128/mbio.03251-21
Copper Induces Protein Aggregation, a Toxic Process Compensated by Molecular Chaperones
Abstract
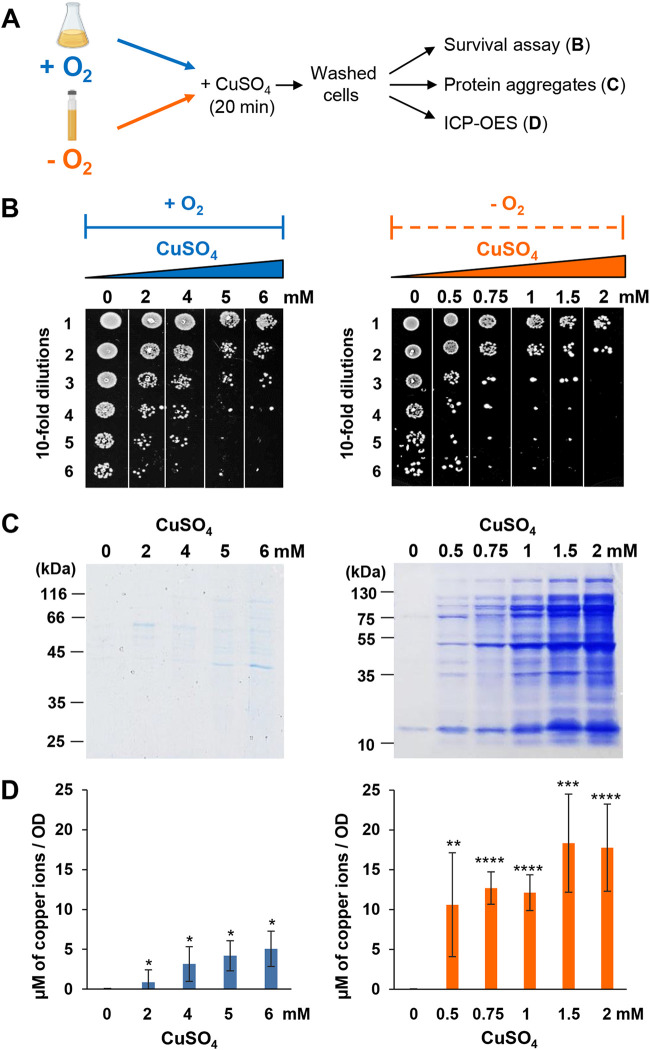
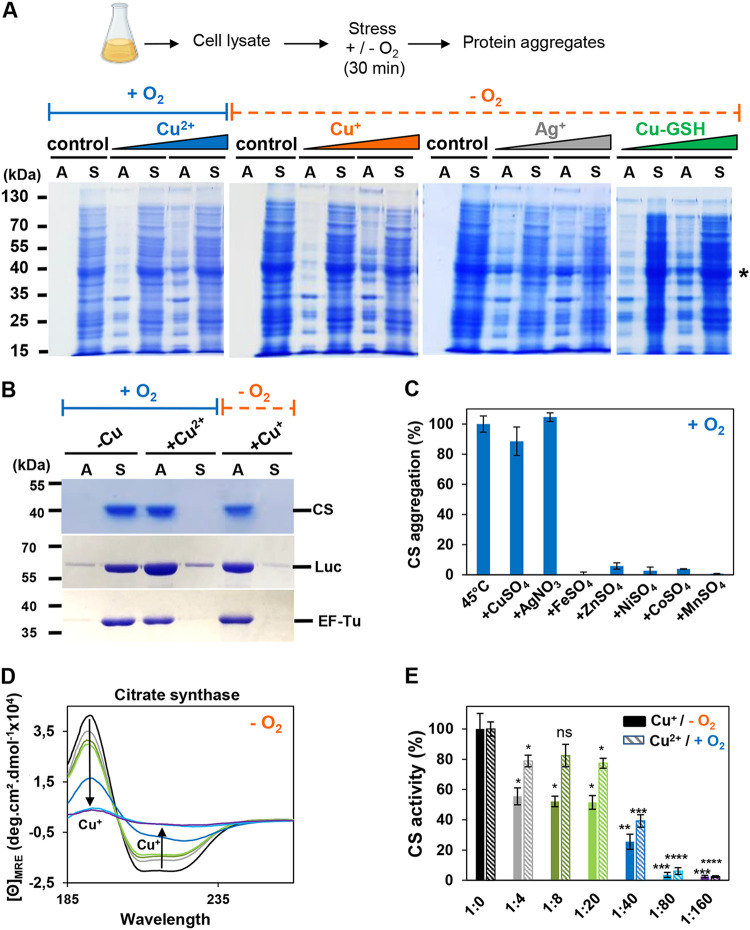
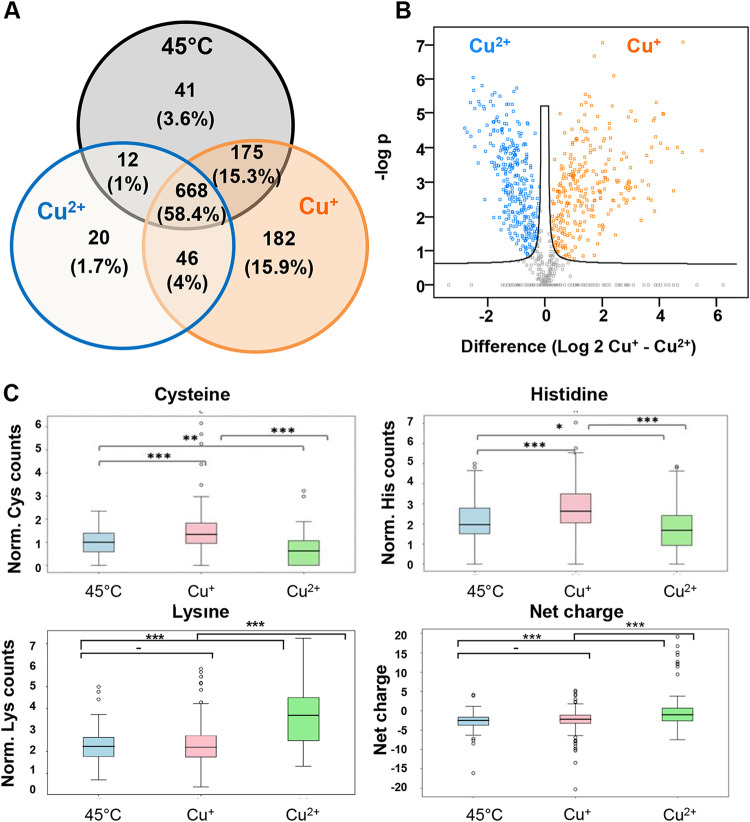
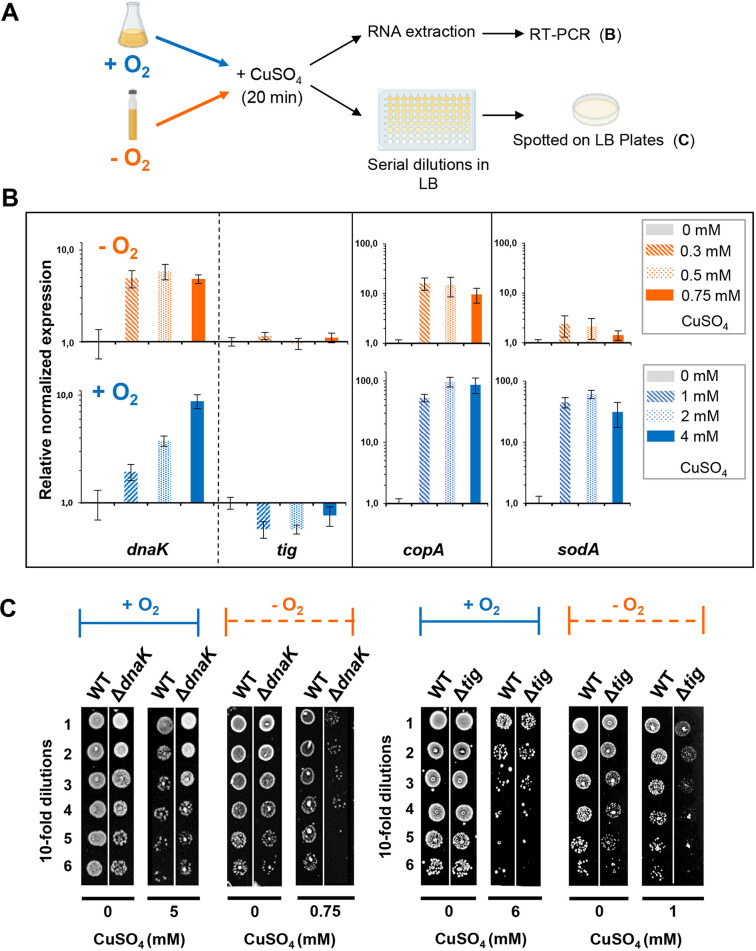
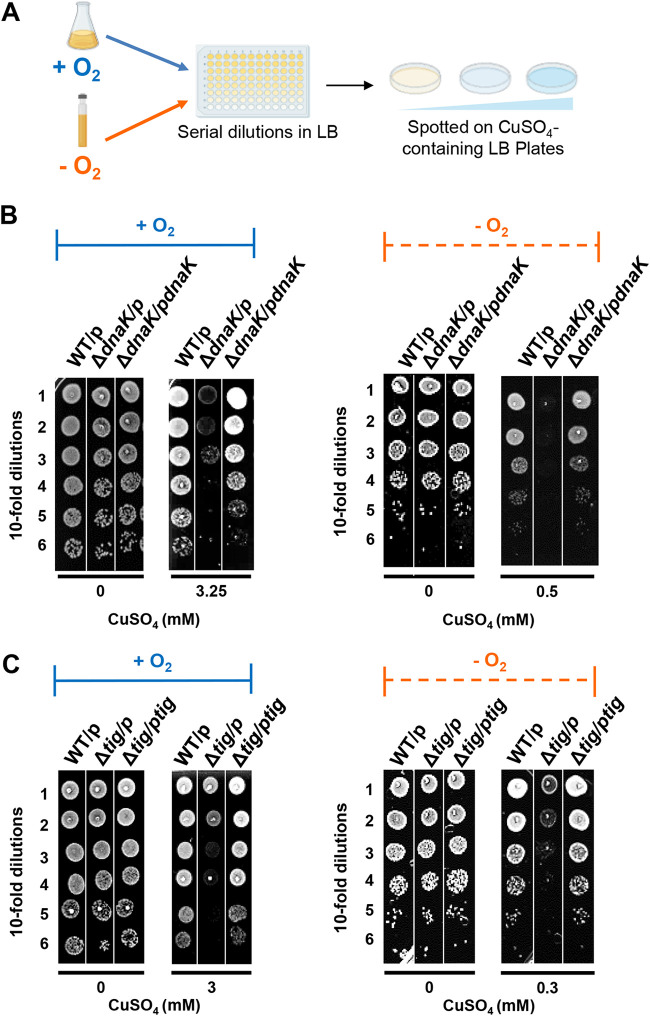
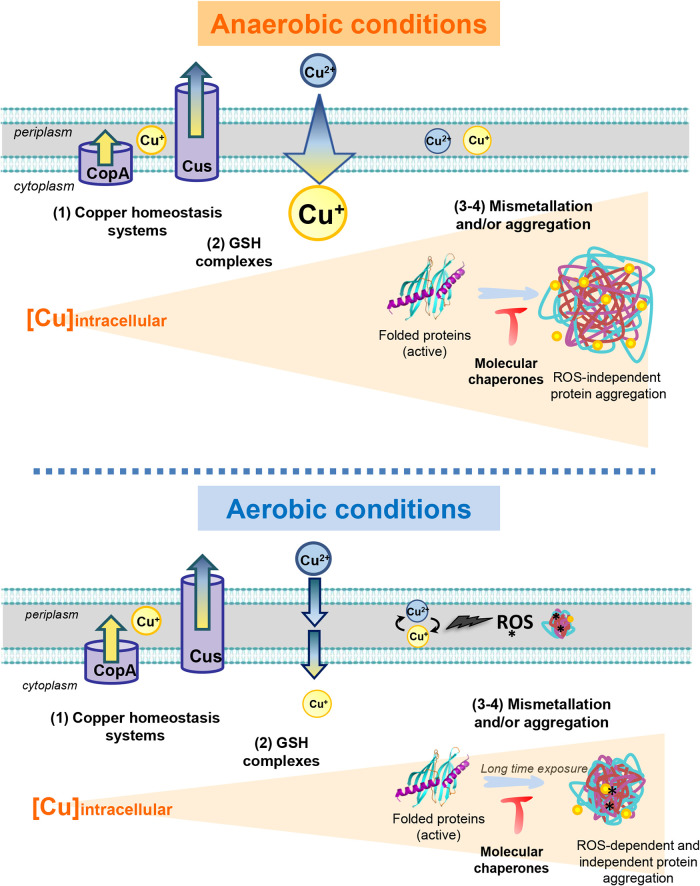
Copper is well known for its antimicrobial and antiviral properties. Under aerobic conditions, copper toxicity relies in part on the production of reactive oxygen species (ROS), especially in the periplasmic compartment. However, copper is significantly more toxic under anaerobic conditions, in which ROS cannot be produced. This toxicity has been proposed to arise from the inactivation of proteins through mismetallations. Here, using the bacterium Escherichia coli, we discovered that copper treatment under anaerobic conditions leads to a significant increase in protein aggregation. In vitro experiments using E. coli lysates and tightly controlled redox conditions confirmed that treatment with Cu+ under anaerobic conditions leads to severe ROS-independent protein aggregation. Proteomic analysis of aggregated proteins revealed an enrichment of cysteine- and histidine-containing proteins in the Cu+-treated samples, suggesting that nonspecific interactions of Cu+ with these residues are likely responsible for the observed protein aggregation. In addition, E. coli strains lacking the cytosolic chaperone DnaK or trigger factor are highly sensitive to copper stress. These results reveal that bacteria rely on these chaperone systems to protect themselves against Cu-mediated protein aggregation and further support our finding that Cu toxicity is related to Cu-induced protein aggregation. Overall, our work provides new insights into the mechanism of Cu toxicity and the defense mechanisms that bacteria employ to survive. IMPORTANCE With the increase of antibiotic drug resistance, alternative antibacterial treatment strategies are needed. Copper is a well-known antimicrobial and antiviral agent; however, the underlying molecular mechanisms by which copper causes cell death are not yet fully understood. Herein, we report the finding that Cu+, the physiologically relevant copper species in bacteria, causes widespread protein aggregation. We demonstrate that the molecular chaperones DnaK and trigger factor protect bacteria against Cu-induced cell death, highlighting, for the first time, the central role of these chaperones under Cu+ stress. Our studies reveal Cu-induced protein aggregation to be a central mechanism of Cu toxicity, a finding that will serve to guide future mechanistic studies and drug development.
Keywords: DnaK; Escherichia coli; copper homeostasis; copper stress; copper tolerance; heat shock; molecular chaperone; protein aggregation; proteostasis; stress response; trigger factor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Comment in
-
Copper Cytotoxicity: Cellular Casualties of Noncognate Coordination Chemistry.mBio. 2022 Jun 28;13(3):e0043422. doi: 10.1128/mbio.00434-22. Epub 2022 May 23. mBio. 2022. PMID: 35604220 Free PMC article.
References
Publication types
MeSH terms
Substances
Grants and funding
- ANR-17-EURE-0003/Agence Nationale de la Recherche (ANR)
- 1537/18/Israel Science Foundation (ISF)
- GM122506/HHS | NIH | National Institute of General Medical Sciences (NIGMS)
- Conseil Régional Provence-Alpes-Côte d'Azur (Région Provence-Alpes-Côte d'Azur)
- ANR-19-CE44-0018/Agence Nationale de la Recherche (ANR)
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
