

A Novel Antimicrobial Peptide Derived from Bony Fish IFN1 Exerts Potent Antimicrobial and Anti-Inflammatory Activity in Mammals
- PMID: 35289673
- PMCID: PMC9045357
- DOI: 10.1128/spectrum.02013-21
A Novel Antimicrobial Peptide Derived from Bony Fish IFN1 Exerts Potent Antimicrobial and Anti-Inflammatory Activity in Mammals
Abstract
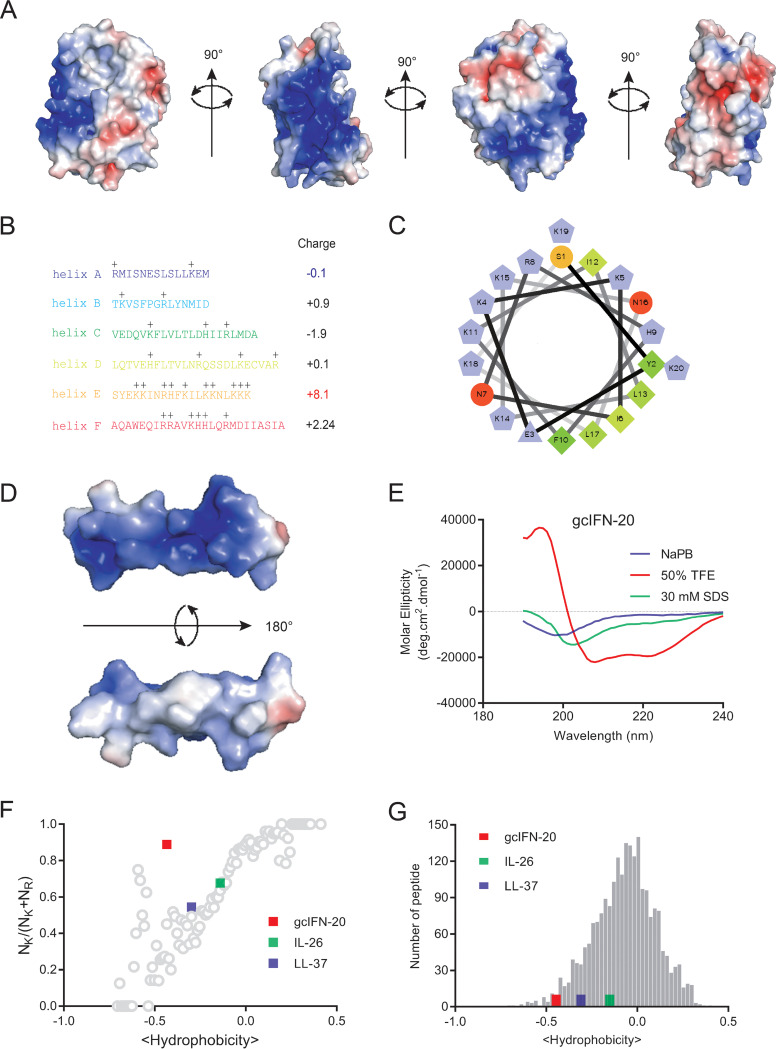
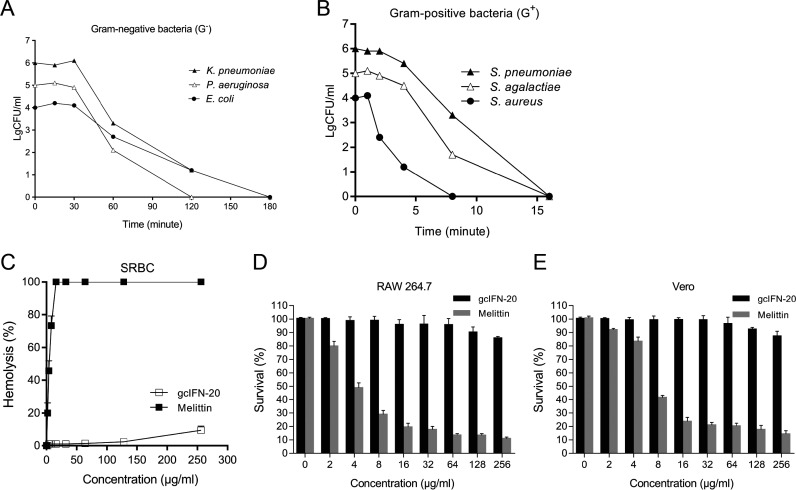
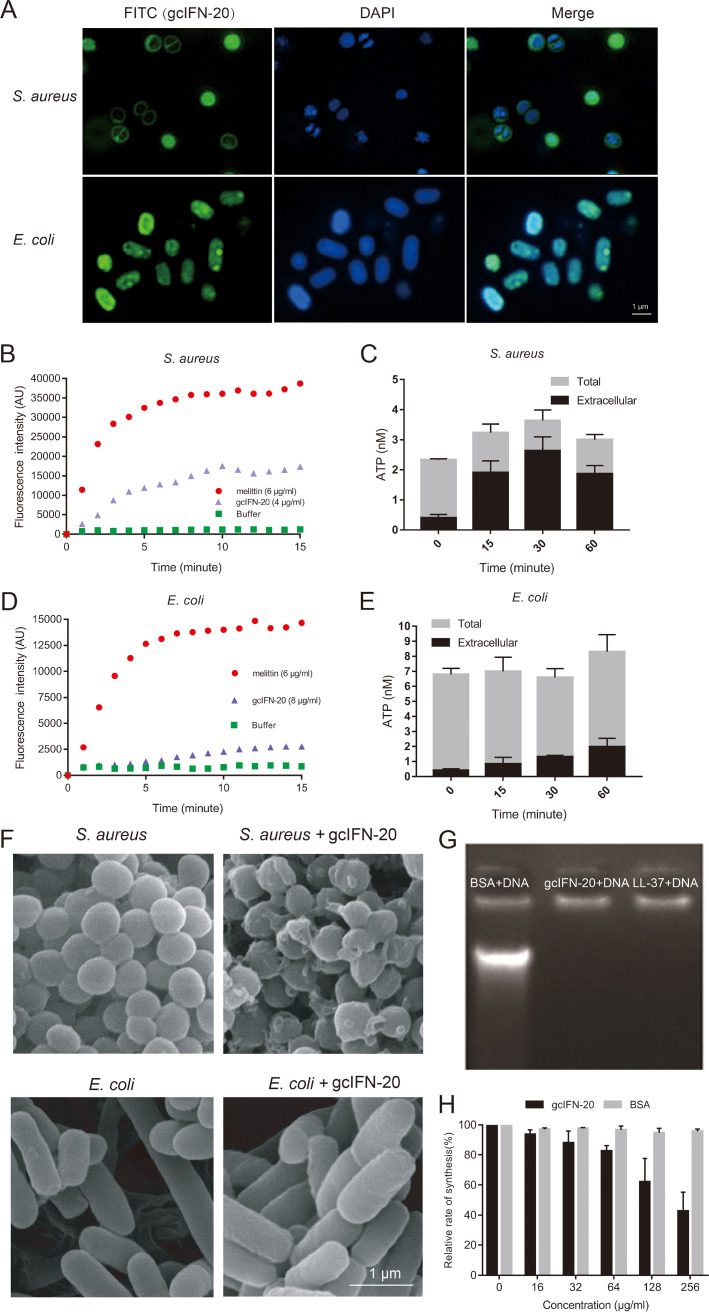
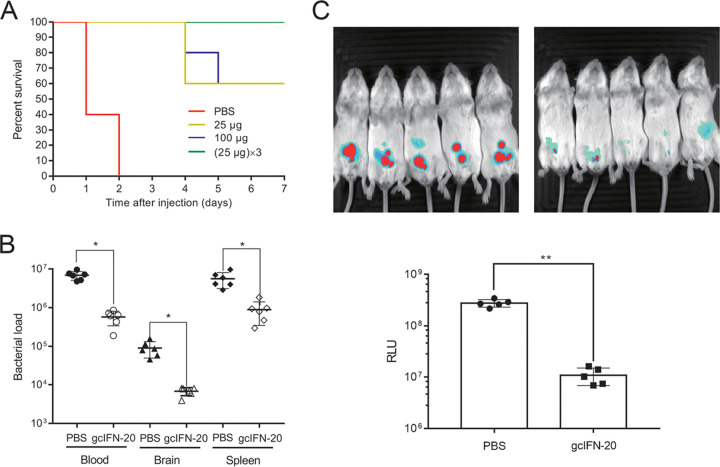
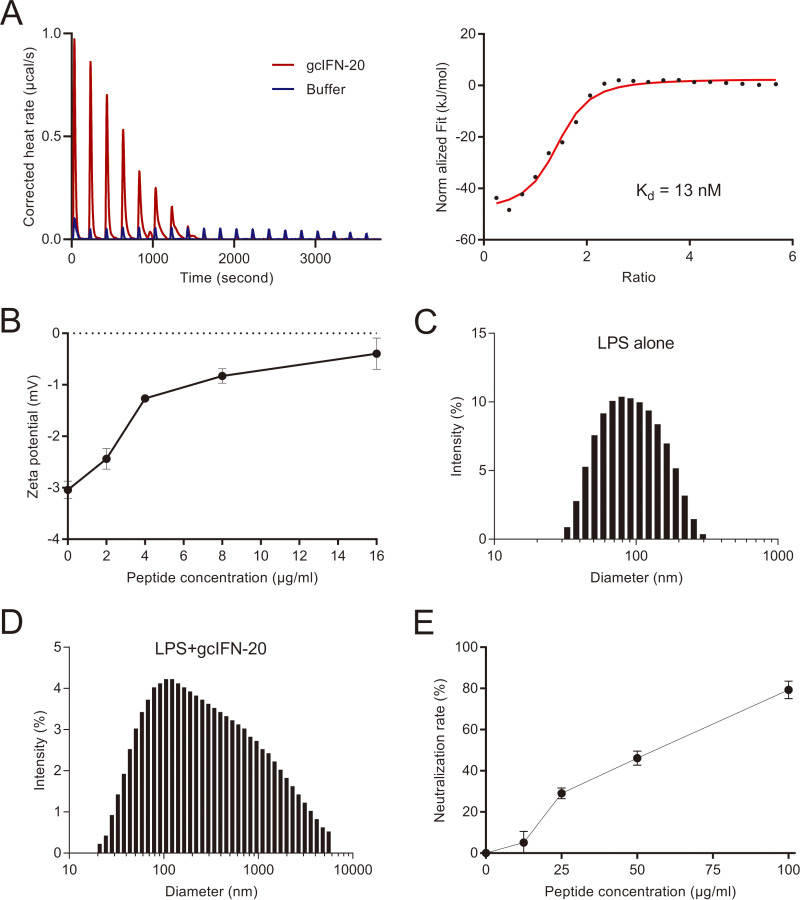
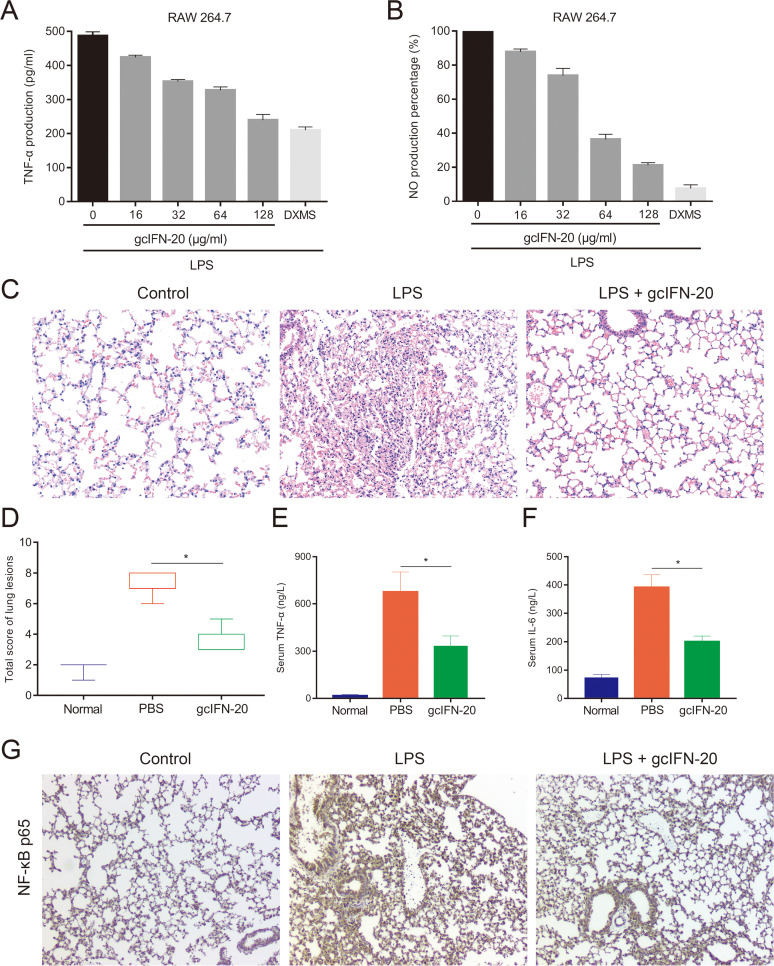
Type I interferons (IFN-Is) are critical antiviral cytokine in innate immunity but with limited direct defense ability against bacterial infections in mammals. In bony fish, despite all the IFN-Is (IFN1-4) act in antiviral immunity, studies demonstrate that IFN1 can remarkably contribute to host defense against bacterial infections. In this study, we found that IFN1 from grass carp (Ctenopharyngodon idella) contains an unusual cationic and amphipathic α-helical region (named as gcIFN-20, sequence: SYEKKINRHFKILKKNLKKK). The synthesized peptide gcIFN-20 could form α-helical structure in a membrane environment and exerts potent antimicrobial activity against multiple species of Gram-negative (G-) and Gram-positive (G+) bacteria with negligible toxicity. Mechanism studies showed gcIFN-20 kills G+ bacteria through membrane disruption and cytoplasm outflow while G- bacteria through membrane permeation and protein synthesis inhibition. In two mouse bacterial infection models, gcIFN-20 therapy could significantly reduce tissue bacterial loads and mortalities. In addition to the direct antibacterial activity, we also found that gcIFN-20 could significantly suppress the lipopolysaccharide (LPS)-induced pro-inflammatory cytokines in vitro and in vivo, obviously alleviated lung lesions in a mouse endotoxemia model. The mechanism is that gcIFN-20 interacts with LPS, causes LPS aggregation and neutralization. The antimicrobial and anti-inflammatory activities in vivo of gcIFN-20 in mammalian models suggested a promising agent for developing peptide-based antibacterial therapy. IMPORTANCE Type I interferons play crucial role in antiviral immunity in both vertebrates and invertebrates. The powerful antimicrobial activity is recently reported in nonmammalian vertebrates. The present study identified a novel antimicrobial peptide (gcIFN-20) derived from grass carp interferon 1, found gcIFN-20 exhibits forceful bactericidal and anti-inflammatory activity in mammals, and efficient therapeutic effect against two clinical severe extraintestinal pathogenic Escherichia coli and a mouse endotoxemia models. The antimicrobial mechanisms are membrane disruption and cytoplasm overflow for Gram-positive bacteria, while membrane permeation and protein synthesis inhibition for Gram-negative bacteria. The anti-inflammatory mechanisms can be aggregating and neutralizing lipopolysaccharide to attenuate the binding with receptors and facilitate phagocytosis. The results indicate that gcIFN-20 can be a promising novel therapeutic agent for bacterial diseases and inflammatory disorders, especially as a potential weapon for multidrug resistant strain infections.
Keywords: anti-inflammation; antimicrobial peptide; bactericidal activity; gcIFN-20; lipopolysaccharide neutralization.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Magana M, Pushpanathan M, Santos AL, Leanse L, Fernandez M, Ioannidis A, Giulianotti MA, Apidianakis Y, Bradfute S, Ferguson AL, Cherkasov A, Seleem MN, Pinilla C, de la Fuente-Nunez C, Lazaridis T, Dai T, Houghten RA, Hancock REW, Tegos GP. 2020. The value of antimicrobial peptides in the age of resistance. Lancet Infect Dis 20:e216–e230. doi: 10.1016/S1473-3099(20)30327-3. - DOI - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Medical
Molecular Biology Databases
Research Materials
