

Octopamine drives honeybee thermogenesis
- PMID: 35289743
- PMCID: PMC8923666
- DOI: 10.7554/eLife.74334
Octopamine drives honeybee thermogenesis
Abstract
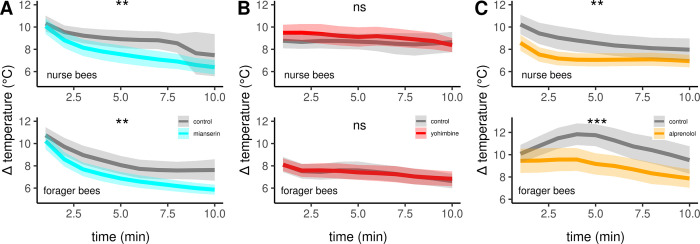
In times of environmental change species have two options to survive: they either relocate to a new habitat or they adapt to the altered environment. Adaptation requires physiological plasticity and provides a selection benefit. In this regard, the Western honeybee (Apis mellifera) protrudes with its thermoregulatory capabilities, which enables a nearly worldwide distribution. Especially in the cold, shivering thermogenesis enables foraging as well as proper brood development and thus survival. In this study, we present octopamine signaling as a neurochemical prerequisite for honeybee thermogenesis: we were able to induce hypothermia by depleting octopamine in the flight muscles. Additionally, we could restore the ability to increase body temperature by administering octopamine. Thus, we conclude that octopamine signaling in the flight muscles is necessary for thermogenesis. Moreover, we show that these effects are mediated by β octopamine receptors. The significance of our results is highlighted by the fact the respective receptor genes underlie enormous selective pressure due to adaptation to cold climates. Finally, octopamine signaling in the service of thermogenesis might be a key strategy to survive in a changing environment.
Keywords: Apis mellifera; GPCR; glycolysis; honeybee; neuroscience; octopamine; thermogenesis.
© 2022, Kaya-Zeeb et al.
Conflict of interest statement
SK, LE, MS, JB, HB, RS, OS, MT No competing interests declared
Figures











References
-
- Åblad B, Ervik M, Hallgren J, Johnsson G, Sölvell L. Pharmacological effects and serum levels of orally administered alprenolol in man. European Journal of Clinical Pharmacology. 1972;5:44–52. doi: 10.1007/BF00560895. - DOI
-
- Baracchi D, Cusseau G, Pradella D, Turillazzi S. Defence reactions of Apis mellifera ligustica against attacks from the european hornet vespa crabro. Ethology, Ecology & Evolution. 2010;22:281–294.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
